CA-SCR-35 is prehistoric site which lies along the northern Monterey Bay coastline, 4.8 km south of the town of Davenport, on a parcel of land known as Edwards Beach. The site is rich in vertebrate faunal remains, and can yield a great deal of information about the early human inhabitants of northern Monterey Bay, the Costanoans** or their immediate predecessors. These remains help to elucidate their subsistence behavior. What animal species are found at the site? Are they primarily marine or land mammals? How were the animals processed by the site's prehistoric occupants? Answers to these questions contribute to answering equally intriguing questions about this little known aboriginal group. Did they perform various animal processing activities in specific areas of the site? Does it appear that the site's occupants shared similar subsistence activities with other aboriginal California groups? Did they occupy the site seasonally or year-round?
To answer these questions my thesis will first outline the site's history as background to the analysis; I will then briefly discuss the site's physical structure and present my research methods. A description of the faunal data will follow, with little extended discussion of their meaning. In this section I will examine the assemblage composition in terms of animal species, the frequency of anatomical elements within each of these groups, and the types and locations of modifications found on the bones. I will also suggest that various agents have modified the vertebrate remains and have produced the assemblage as it is seen today. The following section deals with seasonal or more long-term occupation of the site, and is supported with the natural histories of animals whose remains were identified in the assemblage. In subsequent sections I will use ethnographic accounts of Coast Miwok, Wappo, Coast Yuki, and Tolowa subsistence activities as they relate to animal resources to better understand data from the CA-SCR-35 faunal remains. In this manner, I will present a tentative reconstruction of the subsistence activities at and around CA-SCR-35.
History of the Site
Because the more recent occupational history of the site, and the history of archaeological investigations at the site were presented in detail in the analysis of the assemblage from CA-SCR-35 (Gifford and Marshall 1984), they will simply be summarized here.
The parcel of land on which the Edwards Beach site is located was once part of Rancho El Refugio and belonged to the Mexican government. In 1839 this land was granted to the daughters of a disabled soldier, Jose Joaquin Castro. One of them, Candida Castro, married a Russian sailor, Jose Balcoff; he was officially given the title to the land in 1841. The property was then granted to Bolcoff's sons in 1860; they later returned a portion of the land to their aunt (Mar�a de Los Angeles Castro) who had married Joseph L. Majors.
By 1878 Santa Cruz had become a fairly affluent city whose wealth extended north along the coastline, where dairies were widely distributed. That year Mar�a de Los Angeles Castro Majors sold her parcel of land to Antone and Maggie Silva who established the 40-cow Eagle Glen Dairy. It is quite possible that during the early 1900s the site was used as a source for road surfacing materials, lime, and chicken feed supplement (Gordon 1979:39). The land was to be sold once again to a couple from Switzerland, Mr. and Mrs. Victor Scaroni; they continued to operate the dairy and diversified it in the 1930s by raising pigs on the property.
In 1950, CA-SCR-35 was officially recorded for the first time. Reuben Edwards purchased the property in 1965, his two sons were each given 1/3 interest on the land, which has since been leased out for Brussels sprout farming and as a private beach.
Professor Gordon O'Bannon, of Cabrillo Community College, was in 1967 the first to excavate the site. In 1971 Mr. Ralph Edwards encountered a burial while installing sewage lines on his share of the land. Dr. John Fritz, archaeologist at the University of California at Santa Cruz (UCSC), directed a salvage excavation of three trenches in February of that year and recovered material which was stored at UCSC.
Another excavation conducted in the Spring quarter 1971 in which UCSC, San Jose State University (SJSU), and Harbor High School participated, recovering more material that was taken to SJSU by Dr. K. Bruhns and to UCSC by Dr. Fritz. The latter subsequently loaned some material to West Valley Community College for teaching purposes, but no official record was made of this loan.
In 1975 restricted development of a campground and beach facility on the Edwards property proceeded under archaeological monitoring by a UCSC undergraduate who also acted as site recorder at Dr. Fritz's request.
Before his departure from UCSC in 1976, Dr. Fritz attempted to retrieve the site materials stored at SJSU. He was unable to recover more than a few bags, since Dr. Bruhns had left the school and had not informed her replacement as to the whereabouts of the materials. In 1978 Dr. Diane Gifford, Dr. Fritz's successor at UCSC, laboratory assistant Francine Marshall, and a group of archaeology students analyzed the available CA-SCR-35 materials. When the analysis was nearly completed one student recovered the materials, including some bone remains, loaned to West Valley College. Dr. Gifford and another student, while using the comparative skeletal collection at the Museum of Birds and Mammals at SJSU, were given a substantial number of bag lots of bone from CA-SCR-35. These remains had been stored in the Museum by former curator, Dr. Victor Morejohn. Dr. Gifford and the two students who analyzed the vertebrate remains bagged and gave each bone an artifact record card on which was written a provenience, where applicable, and on some occasions a tentative identification. The bones were stored in the UCSC archives, as it was too late in the quarter to begin any analysis of this new material.
Site Structure
During the 1979 analysis, assessment of the site materials and their proveniences confirmed what Dr. Fritz had originally observed on the basis of surface inspection--the site's constituents were "segregated and clustered spacially" (Gifford and Marshall 1984). Recall the more recent history of the site: the land was used as a dairy and hog farm in the 1930s; cattle trampling and hog rooting behavior (Hafez 1962:339), in addition to mining for road surfacing material and small-scale construction most certainly disturbed the uppermost strata of the site. Ultimately, this would result in a "sugar-bowl" effect on buried materials; smaller pieces are pushed to the lower strata while larger ones rise to the surface (Villa 1982:279). Pocket gophers, which are common in the assemblage, are known for disturbing site stratigraphy by creating networking burrow systems; in doing so they may move enormous amounts of soil to the surface each year (Ingles 1965:211). It is probable that the unmodified pocket gopher bones were secondarily deposited and are the remains of animals who more recently occupied the midden. Stratigraphic proveniences of hydration dated obsidian flakes, plotted on a block diagram of the site, show that the midden is indeed vertically mixed. As a result, one must treat the faunal assemblage as an aggregate that probably spans a considerable period.
Identification Methods
Initially, I identified the bones retrieved late in the Spring of 1979 by using the UCSC comparative skeletal collection in Kerr Hall in conjunction with osteological references by Gilbert (1980) and Brown and Gustafson (1979). The pinniped bones were taken to UCSC Long Marine Laboratory, where an articulated Northern fur seal skeleton is located, as well as complete skulls of local male and female pinnipeds. I concluded that a larger collection was needed, and took the less fragmented, more potentially identifiable bones to the Museum of Birds and Mammals at SJSU.
Levels of identification followed taxonomic hierarchy: class, order, family, genus, and species. Many mammalian bones were fragmented and therefore only identifiable to the ordinal level; it was obvious in these cases that certain anatomically identifiable fragments once belonged to animals which differed greatly in size. Therefore, an arbitrary mammal size classification was developed based on tables of dressed weights of game (Beals and Hester 1974:130-132; see Table 1). Admittedly, weights in this table are idealized, as animal weights fluctuate depending on the season in which they are caught and/or killed. However, other sources on meat weights and animal sizes were unavailable.
Table 1. Mammal Size Classifications
| Size | Weight | Type Animal |
| 1 Very Small Mammal | 1-10 lbs. (0.5-4.5 kg) | Rodent-Gray Fox |
| 2 Small Mammal | 10-50 lbs. (4.5-23 kg) | Grey Fox-Beaver |
| 3 Medium Mammal | <50-100 lbs. (23-45.5 kg) | Beaver-Antelope |
| 4 Large Mammal | 100-500 lbs. (45.5-227 kg) | Antelope-Sea Lion |
| 5 Very Large Mammal | 500-1000 lbs. (227-454 kg) | Sea Lion-Grizzly |
| 6 Very Very Large Mammal | >1000 lbs. (>454 kg) | Grey Whale |
Bones, primarily pinniped and fish that could not be identified at the Museum of Birds and Mammals, were taken to the Departments of Birds and Mammals, and Department of Ichthyology at the California Academy of Sciences in San Francisco. Dr. Jacqueline Schonewald's help with pinniped identifications was invaluable, as was that of Drs. William Follett and Stuart Poss, who were able to identify the less fragmented fish vertebrae. Some of the remains, primarily pinniped, that had been identified in 1979 were also taken to the California Academy of Sciences to recheck identifications. As a result of several taxonomic changes and the new faunal material, vertebrate faunal lists in this paper supersede those of the 1984 publication.
Archaeologists employ various methods to quantify vertebrate assemblages; some methods are more appropriate than others, depending upon individual assemblage characteristics (Grayson 1978). Minimum numbers of individuals (MNI) is an appropriate statistic for calculating faunal abundances at CA-SCR-35 for two reasons. First, this method accounts for element interdependence within species. Second, since the entire site is treated as a single aggregation unit, the potential problems pointed out by Grayson (1978:203-213), which might have arisen as a result of differential aggregation methods, are avoided.
Broad age classes have been established for the identified animal species, where applicable, on the basis of epiphyseal fusion. The rate of fusion is not uniform for every element within a single animal, and also differs between species; therefore, age classes are recognized as immature vs. mature.
THE FAUNAL REMAINS AND THEIR BASIC DATA
Species Present at the Site
Approximately 47% of the pieces in the assemblage are nonidentifiable fragments that are attributed to the vertebrate sub-phylum, or to the mammal, bird, or fish classes. Another 37% is only anatomically identifiable to the class level; 49% of these remains belong to the Large Mammal size category. Bones identifiable to the order and family levels respectively comprise 5.2% and 1.6% of the assemblage. Finally, 9.5% of the assemblage is identifiable to the genus or species level.
The faunal remains at CA-SCR-35 comprise a fairly representative sample of the vertebrate taxa present in the Santa Cruz mountains, and the local (Santa Cruz-Año Nuevo) coastal marsh, nearshore, and offshore regions 1000 years ago (Rudd 1948) with the exception of Chiroptera (bats) and reptiles. Individual taxonomic abundances (Table 2) at the site should in no way be considered a reflection of their actual abundance in the environment at the time of occupation. Numeric representations of taxa are often biased due to differential preservation, nonbiological and biological selective transport of certain animals (or parts of animals) over others, and the methods used to calculate numbers of individual taxa at a site (Gifford 1981; Shipman 1981; Grayson 1978).
Table 2 shows that rodent species were by far the most abundant at the site; they were followed by pinniped and artiodactyl species, in approximately equal quantities, and then by leporid species. The remaining taxa were minimally represented by one to two individuals.
Table 2. MNI of Identifiable Vertebrate Species.
| Taxon | # Pieces
/Taxon | MNI | Age Class | # Pieces/
Age Class |
| Mammals: |
| Scapanus latimanus (Coast mole) | 2 | 1 | AD | 2 |
| Homo sapiens sapiens | 14 | 2 | IMM, AD | 14 |
| Canis sp. (Dog indet.) | 7 | 1 | AD | 7 |
| Canis latrans (Coyote) | 1 | 1 | AD | 1 |
| Vulpes vulpes (Red fox) | 1 | 1 | AD | 1 |
| Urocyon cinereoargentus (Gray fox) | 3 | 1 | AD | 1 |
| Felis sp. (Small cat) | 10 | 2 | IMM, AD | 10 |
| Felis concolor (Mountain Lion) | 2 | 1 | AD | 2 |
| Lynx rufus (Bobcat) | 2 | 1 | AD | 2 |
| Enhydra lutris (Southern sea otter) | 11 | 5 | 2 IMM, 3 AD | 8 |
| Gulo sp. (Wolverine) | 1 | 1 | AD | 1 |
| Mephitis mephitis (Striped skunk) | 3 | 2 | AD | 3 |
| Ursus arctos (Grizzly bear) | 1 | 1 | AG | 1 |
| Eumetopias jubatus (Stellar sea lion) | 3 | 1 | AD | 3 |
| Zalophus californianus (California sea lion) | 28 | 2 | IMM, AD | 25 |
| Arctocephaline (Fur seals) | 21 | 4 | 2 IMM, 2 AD | 21 |
| Callorhinus ursinus (Northern fur seal) | 37 | 4 | 3 IMM, 1 AD | 32 |
| Phoca sp. (True seals) | 10 | 2 | 1 IMM, AD | 7 |
| Phoca vitulina (Harbor seal) | 5 | 1 | AD | 5 |
| Cervus elephus nannodes (Tule elk) | 21 | 3 | 2 IMM, 1 AD | 17 |
| Odocoileus hemionus (Coast mule deer) | 138 | 7 | 3 IMM, 4 AD | 108 |
| Antilocapra americana (Pronghorn antelope) | 2 | 1 | AD | 2 |
| Bos taurus (Domestic cattle) | 10 | 1 | AD | 10 |
| Caprine (Sheep/goat indet.) | 6 | 1 | AD | 5 |
| Lagenorhyncus obliquidens (Pacific white-sided dolphin) | 1 | 1 | AD | 1 |
| Sciurus niger (cf. griseus) (Fox/gray squirrel) | 4 | 1 | AD | 2 |
| Spermophilus beecheyi (California ground squirrel) | 2 | 1 | AD | 1 |
| Neotoma fuscipes (Dusky-footed wood rat) | 1 | 1 | N/A | --- |
| Peromyscus sp. (California mouse, sp. indet.) | 1 | 1 | N/A | --- |
| Microtus californicus (California vole) | 14 | 7 | 1 IMM, 6 AD | 14 |
| Thomomys bottae (Botta's pocket gopher) | 103 | 12 | 5 IMM, 7 AD | 77 |
| Lepus californicus (Black-tailed jack rabbit) | 21 | 5 | AD | 20 |
| Sylvilagus sp. | 29 | 4 | 1 IMM, 3 AD | 22 |
| Sylvilagus bachmani (Brush rabbit) | 27 | 3 | AD | 27 |
| Birds: |
| Gallus sp. | 8 | 2 | N/A | --- |
| Gallus domesticus (Domestic chicken) | 6 | 1 | AD | 6 |
| Butorides virescens (Green heron) | 2 | 1 | AD | 2 |
| Aythya sp. (Canvasback duck) | 1 | 1 | AD | 1 |
| Melanitta sp. (Scoter sp. indet.) | 5 | 2 | N/A | --- |
| Melanitta perspicillita (Surf scoter) | 1 | 1 | AD | 1 |
| Anas sp. (Teal/duck sp. indet.) | 5 | 1 | AD | 4 |
| Anserine (Geese indet.) | 2 | 1 | AD | 1 |
| Chen sp. (Snow goose cf. Ross) | 1 | 1 | AD | 1 |
| Branta canadensis (Lesser Canadian goose) | 4 | 2 | AD | 4 |
| Larus sp. (Gull sp.) | 5 | 1 | AD | 2 |
| Sterna sp. (Tern sp.) | 5 | 1 | N/A | --- |
| Uria sp. (Murre sp.) | 7 | 1 | N/A | --- |
| Uria aalge (Common murre) | 2 | 1 | AD | 2 |
| Ptychoramphus aleutica (Cassin's auklet) | 1 | 1 | AD | 1 |
| Buteo jamaicensis (Red-tailed hawk) | 3 | 1 | N/A | --- |
| Circus cyanus (Marsh hawk) | 1 | 1 | AD | 1 |
| Aquila chrysaetos (Golden eagle) | 2 | 1 | N/A | --- |
| Phalacrocorax sp. (Cormorant sp.) | 9 | 1 | AD | 1 |
| Phalocrocorax auritus (Double-crested cormorant) | 5 | 1 | AD | 5 |
| Gavia sp. (Loon sp.) | 6 | 2 | AD | 4 |
| Tyto alba (American barn owl) | 1 | 1 | N/A | --- |
| Fulmarus sp. (Fulmar sp. indet.) | 1 | 1 | N/A | --- |
| Fulmarus glacialis (Arctic fulmar) | 4 | 2 | AD | 4 |
| Aechmophorus occidentalis (Western grebe) | 1 | 1 | N/A | --- |
| Fish: |
| Rhacochilus toxotes (Rubberlip surfperch) | 1 | 1 | N/A | --- |
| Raja sp. (Skate sp.) | 5 | 1 | N/A | --- |
| Onchoryncus tsawytscha (King salmon) | 1 | 1 | N/A | --- |
| Sardonops sagax (Pacific sardine) | 1 | 1 | N/A | --- |
| Pleuronectidae (cf. Right-eyed flounder) | 1 | 1 | N/A | --- |
| Atherinopsis californiensis (Jacksmelt) | 45 | 1 | N/A | --- |
| GMT | 112 |
Table 2 only includes additional human remains identified since the last analysis was completed. Dr. Lynda Brunker examined the remains in 1979, and at that time the human MNI=9; the two individuals identified in this analysis are most certainly accounted for in the previous MNI. (GMT does not include Homo.)
When the representation of elements within each animal group is compared to its respective MNI, 24 out of 58 mammal and bird species are predominantly, or sometimes solely, represented by long bones or long bone fragments. Long bones, metapodia, and phalanges overwhelmingly represent the larger mammals, while smaller mammals such as canids, felids, and small rodents are accounted for, in majority, by cranial, mandible, and tooth fragments. Brush rabbits, however, are most commonly represented by long bones. Skull fragments and vertebral centra are diagnostic of the great majority of fish species at the site.
Ages of Species at the Site
Where an age distinction is possible, it is apparent that nearly 2/3 of the assemblage is composed of adult individuals. The majority of immature animals are pocket gophers, deer, elk, fur seals, and sea lions.
Studies of age determination based on the rates of epiphyseal fusion (Lewall and Cowan 1963) have been examined to estimate the age range of deer at CA-SCR-35. Using approximate age of long bone and metapodial epiphyseal fusion one may infer that the seven animals ranged from 1 to 4+ years in age. A more precise method of determining age at death of young animals is based on tooth eruption (McLean 1936). Only one immature mandible fragment has been recovered from the site. A second lower premolar (LP2), which had not erupted, is visible inside the mandible; the deciduous premolar (DLP2) was probably in place when the animal died but was subsequently lost. The forthcoming eruption of LP2 presupposes an earlier emergence of the lower incisors (missing)--an event which occurs when an animal is approximately 1 year old. Therefore, this animal is believed to have been 18 months to 2 years old at death. To my knowledge, no research has been conducted on the rates of long bone epiphyseal fusion in local pinnipeds. Consequently, there are no means of even roughly estimating how old these individuals were at death.
Types of Modification
One third of all the animal species, both immature and mature, have been modified in some way. Most commonly occurring modifications are rodent and carnivore gnawing, burning, and cutmarks; often a combination of such features may be observed on a single bone. Burning is the most abundant category, followed by cuts, then rodent and carnivore gnawing (Table 3). Note that each modification type is accounted for with the greatest frequency among long bones and ribs. These values, however, are biased in favor of long bones and ribs because such elements (or fragments of them) are the most abundant in the assemblage. It is also apparent that the remains of some animals are more modified than others (Table 4), and that certain anatomical elements have been repeatedly modified, while other elements have received little or no modification.
Table 3. Assemblage Representation of Modified Remains.
| Burned | Cut | Carnivore
Gnawed | Rodent
Gnawed |
| Antler | 2 | Antler | 1 | Manubrium | 1 | Horncore | 1 |
| Cranial | 26 | Mandible w/teeth | 1 | Sacrum | 1 | Mandible w/teeth | 1 |
| Misc. cranial | 12 | Axial (indet.) | 4 | Ribs | 5 | Vertebrae | 3 |
| Mandible | 12 | Girdle | 2 | Long bones | 16 | Sternum | 1 |
| Teeth (indet.) | 4 | Ribs | 12 | Metapodia | 1 | Scapula blade | 1 |
| Vertebrae | 75 | Long bones | 43 | Patella | 1 | Pelvis | 1 |
| Ribs | 227 | Metapodia | 5 | First phalanx | 1 | Ribs | 6 |
| Long bones | 507 | Phalanges | 2 | -- | -- | Long bones | 21 |
| Girdle | 39 | Astragali | 2 | -- | -- | Metapodia | 3 |
| Metapodia | 50 | -- | -- | -- | -- | Calcaneum | 1 |
| Podia | 6 | -- | -- | -- | -- | -- | -- |
| Phalanges | 44 | -- | -- | -- | -- | -- | -- |
| Sternum/furculum | 7 | -- | -- | -- | -- | -- | -- |
| Patella | 3 | -- | -- | -- | -- | -- | -- |
| Astragalus | 2 | -- | -- | -- | -- | -- | -- |
| Calcaneum | 3 | -- | -- | -- | -- | -- | -- |
| Nonidentifiable | 650 | -- | -- | -- | -- | -- | -- |
| | Burned | Cut | Carnivore
Gnawed | Rodent
Gnawed |
| % of modified assemblage | 92.4 | 4.0 | 1.4 | 2.2 |
| % of total assemblage | 19.5 | 0.8 | 0.3 | 0.5 |
| Category weight (g) | 1,514.7 | 884.6 | 380.8 | 541.0 |
Table 4. Modified Identifiable Faunal Remains.
| Taxon | Modification Type | Elements |
| Urocyon cinereoargentus | B | uln
| Felis sp. | B | pha1
| Enhydra lutris | B | man, ang, lbn, hum, tib
| Ursus arctos | CT, R | mant
| Eumetopias jubatus | B, CA | uln, mc0, pat
| Zalophus californianus | B, CT, CA | tth, rad, fem, mt0, fp20, hp20, pel, hum, rad, uln, fem, tib, pha1, pat
| Arctocephaline | B, CT | pel, hum, rad, uln, fem, tib, pha1, pat
| Callorhinus ursinus | B, CT, CA | hum, rad, fem, mc0, mt0, fp10
| Phoca sp. | B, CT | ang, mc0
| Cervus elaphus nannodes | B, CT, CA, R | atl, tho, rib, tib, tibe, met, pha1, pha3, ast
| Odocoileus hemionus | B, CT, CA, R | ant, pet, scpg, hum, rad, fem, tib, tibe, met, mete, mcm, mtm, pha1, pha2, ast, cal
| Antilocapra americana | B, R | hco, tib
| Bos taurus | B, CT (sawed), R | met, rib
| Caprine | B | lum
| Lepus californicus | B | max
| Sylvilagus sp. | B | man, sym
| Sylvilagus bachmani | B | max, fem, tib
| Gallus sp. | R | ste
| Butorides virescens | B | hum
| Melanitta sp. | B | uln
| Anas sp. | B | cor, cmc
| Larus sp. | B | car, fp11
| Buteo jamaicensis | B | hp20
| Aquila chrysaetos | B | hp20
| Phalacrocorax sp. | B | tbt
| Sardonops sagax | B | opr
| Pleuronectidae | B | vrt
| Atherinopsis californiensis | B | vrt |
|
Key:
Modification type: B=burned; CT=cut; CA=carnivore gnawed; R=rodent gnawed.
Elements: max=maxilla; hco=horncore; pet=petrosal; opr=operculum; ant=antler; man=mandible; mant=mandible with teeth; sym=symphysis; ang=angular; tth=tooth indet.; vrt=vertebra indet.; atl=atlas; tho=thoracic vertebra; lum=lumbar vertebra; rib=rib indet.; ste=sternum; scpg=glenoid of scapula; cor=coracoid; pel=pelvis indet.; lbn=longbone indet.; hum=humerus; rad=radius; uln=ulna; met=metapodial indet.; mete=metapodial epiphysis (unfused); mc0=metacarpal digit indet.; mcm=main metacarpal; cmc=carpometacarpus; fem=femur; tib=tibia; tibe= tibia epiphysis (unfused); tbt=tibiotarsus; mt0=metatarsal digit indet.; mtm=main metatarsal; pat=patella; car=carpal; ast=astragalus; cal=calcaneum; pha1=phalanx 1; pha2=phalanx 2; pha3=phalanx 3; fp10=front 1st phalanx digit indet.; fp11=front 1st phalanx 1st digit; fp20=front 2nd phalanx digit indet.; hp20=hind 2nd phalanx digit indet (Gifford and Crader 1977).
|
| | | | | | | | | | | | | | | | | | | | | | | | | | |
Based on the data presented above, one may ascertain that several taphonomic agents have independently and collectively acted upon the CA-SCR-35 fauna, both prior to, and after its deposition. Further examination of the assemblage will reveal that these agents are nonbiological and biological; the latter may be specified as plants (to a very small extent), rodents, carnivores (predator-scavengers), and humans.
Weathering and Chemical Decomposition
Nonbiological agents responsible for the partial destruction of a bone assemblage can include both physical and chemical weathering processes. The extent to which bones physically break down is determined by their microscopic structure (Behrensmeyer 1978:153; Shipman 1981:372), hence the physical condition of the animal at its time of death.
According to Behrensmeyer, the physical and organic decomposition of bone lying on land surfaces is enhanced by fluctuations in temperature and humidity. Regional temperatures in the Monterey Bay area do not appreciably fluctuate diurnally or annually because of offshore winds and deep water upwelling. However, the amount of precipitation can be quite variable from year to year (Gordon 1979:14-15). The degree to which these factors affect the physical cracking and splitting of bone remains in this region is not known. Variations in the amount and intensity of rainfall may contribute more to their destruction than do temperature fluctuations.
Weathering stages (0-5) were established by Behrensmeyer (1978) to help taphonomists determine approximately how long a given assemblage has been exposed to the "elements," that is, how much time has elapsed since the time of death. Although they were established for bones decomposing in a climate much drier than that of the Monterey Bay area, these stages provide a basis for describing the relative condition of bone remains. Bone that shows no cracking or flaking due to weathering is ascribed to weathering Stage 0. Stages 1-2 represent elements which are cracked and flaked, and which may no longer possess the outermost layer of bone. Remains that are in progressively more advanced stages (3-5) of physical and chemical weathering acquire a fibrous texture, are extensively cracked and fragile, and eventually fall apart in situ (cf. Behrensmeyer 1978:151).
Formal weathering stages have not been described for the CA-SCR-35 assemblage because the mechanical disturbances which occurred at the site are believed to have affected the length of time to which the remains were exposed to natural elements. Admittedly, relative weathering stages would have been helpful in determining the overall condition of the remains. After examining the bone assemblage, it is my impression that the overall condition of the mammal remains ranges from Stages 1-3 (Behrensmeyer 1978:151-152). The extent to which the non-animal agents have affected the faunal remains is believed to be of minor importance relative to the affects of animal agents.
It is fair to assume that chemical weathering, often greater at the soil and air interface than below ground, also played a role, albeit negligible, in the decompositional history of the CA-SCR-35 remains (Behrensmeyer 1978:154). Chemically, bone is known to have a high survival rate in midden soils, where the pH is neutral or only slightly alkaline (Halley 1980). Traces of root etchings are present on several pieces of bone suggesting that plant roots modified at least a small portion of the assemblage by secreting various organic acids into the soil (Shipman 1981:375; Behrensmeyer 1978:155; Evans 1978:73). The degree to which other substances, such as cattle excrement and crop fertilizers, chemically affect the bones is not known.
Rodents
Evidence of rodent gnawing activity at the site is quite infrequent (Table 3). Rodents prefer "clean" bones, ones that are free of fat and flesh, and slightly weathered (Gifford 1981:414). Diagnostic rodent gnaws are distinguished as two parallel sets of wide, flat, shallow, grooves which are often present along the broken edges of the elements in question (Shipman 1981:368). Only Large to Very Large (land) Mammal bones are rodent gnawed; none of the pinniped remains exhibit characteristic rodent tooth marks (Table 4). It appears that the rodents at CA-SCR-35 preferentially gnawed the harder artiodactyl remains over those softer, less dense, and perhaps more odoriferous bones of pinnipeds. The artiodactyl bones are undoubtedly better suited for keeping rodent incisor growth in check. Such growth can be extremely rapid; for example, Botta's pocket gophers' incisors may grow 46 inches per year (Ingles 1965:207).
Carnivores
The bone remains show that there has been carnivore activity at the site. Binford has described in detail (1981:35-85) the characteristic features (fractures, tooth marks, punctures, pits, scores, and furrows) which carnivores habitually leave on bones after having gnawed them. The majority of "unequivocal" carnivore gnawing is in the form of pitting, scoring, and furrowing. One half of these gnawed bones are minimally identifiable mammal long bone and rib shaft fragments which have received no other type of modification (one exception is burned). Less identifiable mammal long bone fragments are occasionally spirally fractured, are pitted, or are chipped along the broken edges; rib shaft fragments are typically crenulated along these broken edges. The remaining bones are primarily pinniped and artiodactyl long bones.
Zalophus remains show light to heavy gnawing in the form of scoring and furrowed edges of proximal and distal ends and articulations. One femur is more heavily gnawed than other bones, the head and lesser trochanter have been completely removed (furrowing) but the greater trochanter remains intact. Its shaft is scored, furrowed, and the medullary cavity is exposed along the distal medial side. The distal medial condyle has been removed, while the lateral one is lightly furrowed. Only one specimen (Callorhinus humerus) has been punctured, presumably by a fairly large canine (0.75 cm in diameter). The puncture is located on the distal posterior side, just above the articular surface. No other gnawing feature is present on the bone.
The majority of gnawed artiodactyl remains are hind limb fragments, although a few ribs are scored and have crenulated edges. One tibia (Figure 6a) and three femurs all belonging to Odocoileus exhibit light to moderate carnivore gnawing. The lateral border of the tibia's proximal articulation is furrowed, exposing the cancellous bone, and the patella is missing (furrowed). Two of the three femurs belong to immature individuals; the proximal end of one is completely missing (scored shaft and irregular edges), and the other's distal end is lightly furrowed. The third femur, a proximal fragment, lacks a greater trochanter (furrowed).
Carnivores gnaw bones in a sequential and systematic manner (Haynes 1980); the degree to which each element of a consumed animal has been gnawed reflects the degree of carcass utilization by carnivores. According to Binford's observations of wolf kills, dens, and dog yards, furrowing, puncturing, and crenulated edges of bone predominantly occur at kill sites, while more extreme furrowing, extensive pitting, and scoring are more common features of "boredom chewing" (1981:49). When gnawing bone shafts, carnivores sometimes spirally fracture them, and in cases of intense gnawing, wolves and Eskimo dogs typically channel into long bone shafts after having gnawed the softer cancellous bone at the epiphyses (Haynes 1983; Binford 1981:51).
Although the evidence is far from conclusive, the location of the puncture mark on the Callorhinus distal humerus suggests that carnivores differentially treated pinnipeds from other animals as far as initially disarticulating them to obtain choice pieces of meat from fresh carcasses. When dismembering an artiodactyl carcass, carnivores will disarticulate the scapula-proximal humerus joint by eating through it before attacking the lower part of the forelimb (Binford 1981:44). Since this particular element's proximal end is intact (with the exception of several definite cuts), it appears that this animal was not dismembered in such a sequence. Because heavy muscle tissue and fat surrounds pinniped forelimbs, an easier way to go about disarticulating such an animal may be to first attack a less concealed portion of the limb segment.
In general, carnivore gnawed remains from CA-SCR-35 suggest that their activity at the site has been light to moderate; with two exceptions, the bones are not intensively gnawed. Although there are not many species of native carnivores which could have potentially gnawed the bones, one would expect the evidence for carnivore activity at the site to be more substantial (Table 3), given the hydration dates which allow for at least 2,000 years occupation.
Why is carnivore gnawing the least frequently encountered modification? The assemblage representation of anatomical parts does not reflect that of a carnivore kill site in which one would expect to find more remnants of axial/girdle elements than limbs, because the latter would be dragged off the site by hungry animals (Binford 1981:42). Dairy cow trampling may have destroyed the gnawing evidence by crushing buried site materials under hoof. However, even if this were the case at least some evidence of more intense gnawing, such as heavy pitting and channeling, would be present on the bones. It is not. A remaining alternative is that carnivores did not have immediate access to the bones while they were still attractive to chew, and even after they were no longer quite so appealing.
Burning
The majority of modified bones in the assemblage (ca 92%) are burned (Table 3); 40% of these are nonidentifiable bone fragments. This is the only type of modification represented on all anatomical regions; more often than not, burning is light rather than severe. The most frequently encountered identifiable burned specimens are fragments of long bones and ribs belonging to Medium and Large Mammals, with limbs being twice as abundant as ribs. More specific taxonomic identifications reveal that much of the burned bone is artiodactyl and pinniped. Among pinniped remains, the articular ends of long bones, and more extensive regions of metapodia and phalanges, are most often burned; elements are equally represented in this category. Burned artiodactyl remains are more abundant and are represented by approximately three times as many metapodia and phalanges as long bones. In this instance, the ends of long bones and metapodia are burned, while phalanges are more thoroughly burned. Several of these bones, primarily hind limb fragments belonging to pinnipeds and artiodactyls, had been subsequently gnawed by carnivores.
Interestingly, none of the identified rodent remains are burned; only three of the indeterminate rodent bones are burned. Even if one includes the Very Small Mammal category, which includes local rodents, only 0.7% are burned.
Cutmarks
The last type of modification recognized is cuts on the surface of the bones. Cutmarks made by stone tools are distinguished from other types of modifications as series of fine, parallel striations (deeper and narrower than those resulting from rodent gnaws) which lie perpendicular or parallel, and some times oblique to the long axis of the bone (Shipman 1981:365; Potts and Shipman 1981:577; Binford 1981).
The majority of animals represented in this category are minimally identifiable Medium and Large Mammals, pinnipeds and artiodactyls. Long bone shaft fragments belonging to the minimally identifiable mammals possess cuts both perpendicular and transverse to the long axis of the bone. Ribs are cut transversely along the shafts, or in some cases near the proximal articulation.
Cut pinniped remains, few in number, are primarily metapodia and phalanges along with two long bones. Two transverse cuts are present below the head of a Callorhinus humerus on the posterior medial side. A Zalophus radius is also cut along its shaft and distal shaft. For the most part, cuts on metapodia and phalanges lie on the proximal shafts.
Large to Very Large indeterminate artiodactyl ribs have been sawed at the proximal ends, and vertebrae are sawed through their centra. Domestic cattle metapodia are cut distally, just above the articulation; I was unable to determine whether these cuts resulted from metal or stone tools. Tule elk remains exhibit cuts across a thoracic spine, as well as on an astragalus where they lie on its posterior-lateral side. Mule deer remains are more abundant than those of tule elk, hence as a result of sampling bias they appear to be cut with a higher frequency. Transverse cuts are present on one medial distal humerus, just above the articulation. More than six similar cuts are found above and below a femoral lesser trochanter. Similar cuts are located on the anterior and posterior shafts of two other femurs, one of which is carnivore gnawed at its distal end. Cuts on two proximal tibia fragments lie across the tibial crest, near the patella, and along the lateral posterior margin of the articulation. The anterior medial astragalus and lateral metapodial epiphysis also possess cuts. One remaining specimen is worth mentioning in this discussion--a complete California grizzly mandible possesses a series of cuts along the anterior edge of the masseteric fossa and several longer ones within the fossa itself. Additional cuts are found medially, directly below the LM1-2 row.
INFERENCES FROM THE FAUNAL REMAINS
Origins of the Assemblage
In the past, bone assemblages were thought to have been humanly modified because it was assumed that humans are the only taphonomic agent capable of spirally fracturing limb shafts, of leaving "cutmarks" on bones, of removing tuberosities, etc. during the process of butchery (this includes dismemberment, muscle removal, marrow extraction, etc.) and of making "bone tools." More recently (e.g., Shipman 1981; Haynes 1980, 1983; Binford 1981), these assumptions have been refuted by demonstrations that humans are not the sole creating or modifying agents capable of leaving such traces in bone assemblages. Therefore, such conclusions cannot be made without first considering the role of other possible agents.
Morphological examination of modification on the CA-SCR-35 assemblage has established that rodents and carnivores are two agents responsible for modifying the assemblage as we see it today. Remnants of their activities, however, do not explain all of the modifications encountered. Human modification of most of the assemblage is inferred from the presence of burned and cutmarked bone, as well as from bone breakage and damage patterns and overall element representation.
The previous analysis (Gifford and Marshall 1984) established that the Edwards site had been occupied by aboriginal Californians. Five formal chert tools, primarily retouched bifaces, were examined in 1982 by Dr. Nicholas Toth, of the University of California, Berkeley. Dr. Toth, who microscopically distinguished meat and bone polishes on the tips and edges of these tools, concluded that they were hunting and/or butchery paraphernalia. A large number of unretouched flakes was also found at the site; Phillip Walker's experiments have shown that such flakes are more effective than bifaces in processing marine mammal carcasses (Walker in Gifford and Marshall 1984). Clearly, the site's inhabitants had the technology to effectively hunt and process animals for food and other resources; thus, the CA-SCR-35 occupants are believed to have generated most of the vertebrate assemblage.
Season(s) of Site Occupation
Gifford and Marshall (1984) state that invertebrate and vertebrate remains indicated that CA-SCR-35 was inhabited during the winter months, and may even have been a permanent habitation site, as it is in such close proximity to a variety of aboriginally useful vegetation zones. The season of occupation was not expected to change with the addition of new vertebrate faunal material. However, natural history information on the main groups of animals was examined to check agreement between the hypothesis and the faunal assemblage as a whole.
Movement of Indian seasonal encampments has been thoroughly documented ethnographically and will not be discussed in detail here. Suffice it to say that the Costanoans are believed to have moved their villages to the coast during the winter to early spring months (December-March/April) so that a variety of molluscs, other shellfish, and marine resources could be collected; the coastline also provides a much drier climate than the hills, which occasionally receive more than twice the coastal rainfall in winter. Summer and autumn months were spent in the woodland foothills, where the Costanoans could gather acorns, nuts, and other plant foods, and hunt large and small game; the inland climate is warmer and drier than along the coast, where dense fog settles at this time of year (Margolin 1978:13; Gordon 1979:13). Gifford and Marshall (1984) note that since CA-SCR-35 is in close proximity to uplands, a major seasonal residential shift may not have been as necessary as in the Pajaro or San Lorenzo Valleys.
There are both marine and anadromous fish species at CA-SCR-35 (Table 2). Marine species are typically found year-round along the California coast in nearshore or inshore, rocky to soft sand-mud bottoms. Sardines, noted for their oily flesh, are primarily marine, although many move into brackish and fresh waters. The Pacific sardine spawns offshore in spring and summer months, while other sardine species migrate to rivers and spawn in freshwaters. Flounders are also primarily marine, however, the Pacific Starry flounder is found in brackish waters of estuaries and occasionally enters freshwater. The presence of possible eel slitters (Figures 6a and 6b) suggest that lampreys may have been a resource exploited by the inhabitants of CA-SCR-35. These eel-like fish range along the entire length of the North American continent. Adults are usually marine, but enter freshwater streams and lakes in the spring and summer to spawn. The largest anadromous species found at the site is the King salmon. While fall spawning occurs with greater regularity, some salmon spawn in the spring (Baumhoff 1963:174). Today, King salmon are rarely found in freshwater south of the Sacramento-San Joaquin River system (Tesch 1977; Herald 1972; Eschmeyer et al. 1983; see Harrison 1892 as well, for a slightly dated Monterey Bay fish species list).
The bird assemblage is primarily composed of waterfowl and marine species; some of these are only seasonally present in the area, notably several species of ducks and geese (Gifford and Marshall 1984:42). Four new bird species' remains at the site include the Red-tailed hawk, Golden eagle, American barn owl, and the Western grebe. The birds of prey are all commonly found in the oak woodland and grassland ecozones. Western grebes typically winter along the California coast from October to April. Essentially, bird remains do not supplement the seasonal information presented in the 1984 publication. Remains of rodents and Small-Medium Mammal specimens are likewise not useful seasonal indicators because the animals are resident and may be found along the California coast (Ingles 1965).
Pinnipeds are among the more useful indicators of aboriginal seasonal activity. Phoca, arctocephalines, and Callorhinus were identified in 1979; new species include the Stellar and California sea lion. Harbor seals (Phoca) are still commonly found along the California coast, and although they are the wariest of all local pinnipeds, they often haul out onshore or on offshore rocks and are seldom seen at sea (Allen 1974:488; California Coastal Marine Studies 1980:34).
Stellar sea lions (Eumetopias) descend the eastern Pacific coast from the Pribilofs. Females are the first to arrive in northern California in early January, while the species as a whole is most commonly seen off California shores from May to early July. The majority of animals are found on an important rookery, north of Año Nuevo Island (California Coastal Marine Studies 1980:32-33).
California sea lions (Zalophus) are the most abundant pinnipeds along the California coast. In early May they migrate south towards rookeries extending from the Channel Islands down to the west coast of Mexico, however, non-breeding aggregations occur as far north as British Columbia (California Coastal Marine Studies 1980:29; Peterson et al. 1967:4). The most important aggregation of migrating animals was observed in early May on Año Nuevo Island. When not migrating, these animals are infrequently seen at sea. Presently, sea lion populations are believed to be similar to what they were before the species was exploited for furs, after fur seals became scarce (Le Boeuf, personal communication 1984).
The southern, or Guadalupe, fur seal (Arctocephalus) is not known to migrate to northern California waters, but may have in the past. Mid-19th century sightings of Southern fur seals around Monterey may in fact be erroneous, since a distinction between the species had not been made at the time (Starks 1922:159).
The northern fur seal, Callorhinus, breeds and gives birth on the Pribilof Islands from June to September-early November. At the end of the breeding season (early November) the females and newly weaned pups migrate south for the winter. They are found along the California coast as far south as San Miguel Island (permanent hauling ground) from early December to May or June, with population peaking in February (California Coastal Marine Studies 1980:24). Individuals have been observed to come ashore, along the northern California coast, for rests between feedings (California Coastal Marine Studies 1980:25; Le Boeuf, personal communication 1984). Late 19th century accounts of northern fur seal migratory behavior note that the animals left the Pribilofs in December to return in May; this correlates well with their present day migratory habits. However, by the turn of the century, migrating animals had virtually ceased to haul out. Northern fur seals are perhaps returning to "pre-exploitation habits," since as a species, they have been protected by law for nearly a full century and are recovering from over-exploitation. Conclusions of this nature, possibly premature, may not be drawn at the present time (Le Boeuf, personal communication 1984).
The physiologic condition of pinnipeds is best in mid-late spring (March-May) (Le Boeuf, personal communication 1984). Therefore, aboriginal hunters would have benefitted the most from seals and sea lions if they hunted them most intensively in the spring. Based solely on the characteristics of migratory behavior and on the presence of both mature and immature pinnipeds at the site, the time of occupation ranges from late fall to late spring (early December-May).
Once present in coastal northern California in great numbers, deer, elk, and antelope are of limited use in seasonality studies because, although some elk and antelope herds were migratory, those locally present are believed to have been resident. Seasonal antler development in mule deer and elk bucks may be of some use in determining season of death. In young mule deer males antler growth is visible approximately three months after birth. By mid-summer the fawns have small developing pedicels beneath the fur on their frontal bones; small antlers are visible by autumn. Further development of antlers ceases through the winter months and pedicels are covered and furred with skin. In the spring, outward growth of the antlers resumes, and by late summer unbranched antlers mature. In its second year the animal usually produces a pair of forked antlers. Mature bucks shed their antlers from December through February. New growth of antlers resumes in late March and continues into July (Linsdale and Tomich 1953).
As is the case with all wild animals, these artiodactyls are in better physiologic condition during certain times of the year than during others. The aboriginal practice of burning the oak woodland and grasslands in the fall resulted in optimal forage for artiodactyls in spring (Lewis 1973). Judging from the amount of energy invested in hunting, the hunters would want maximum yield for their efforts. In other words, a hunter would have been most likely to seek deer, elk, and antelope when they were in prime condition. This would not have been in late fall through the winter (November-January) when the animals derive less nutrition from browse and other forage (Wallmo 1981; Linsdale and Tomich 1953). Optimal forage is available for wild artiodactyls in the spring and early summer months (Linsdale and Tomich 1953; McCullough 1969; Kerr 1979). Thus, it is conceivable that aboriginal hunters would have most actively sought these animals in mid-spring to late summer/early autumn.
Year Round Habitation of CA-SCR-35
and Meat Input Estimates
The range of time for occupation of CA-SCR-35 spans at least 2,000 years. During this time, the site may only have been periodically occupied. The grand minimum total (GMT) for the vertebrate assemblage is 112 individuals. Using Beals' (1974:130) meat weight chart, the minimum amount of meat represented by only a portion of the GMT is 3,000 lbs. This figure does not include marrow, blubber, fish, or usable meat weights for animals which do not appeal to most modern Californians' epicurean tastes (e.g., mice and sea birds). The amount of usable meat is expected to be closer to 4,500 lbs., especially since I arbitrarily determined that the immature animals possess about half the usable meat than adults of the same species. Overall, 3,000 lbs. of meat is a substantial figure, but given the 2,000 years of occupation time allowed by obsidian hydration dates it is quite small. The data presented above lead one to believe that first, if the site was occupied year-round, it was not continuously occupied, and second, vegetable, shellfish, and perhaps other resources whose remains were not recovered from the site (e.g., reptiles and insects) formed an important part of the CA-SCR-35 aboriginal diet. The total CA-SCR-35 shellfish component includes over 25,000 fragments and weighs approximately 54 kg (Gifford and Marshall 1984:38). It is difficult to determine the amount of meat represented by this figure because MNI are unavailable for the component. Even if they were available, since shellfish grow continuously throughout their lives, immature or mature MNI categories would be irrelevant. At best, one may be able to estimate individual shellfish sizes (and MNIs) based on umbo (beak) fragments, and then approximate the amount of meat for the various sizes of shellfish species present in the assemblage.
Available information suggests that CA-SCR-35 was inhabited during all seasons of the year. Faunal remains and natural history backgrounds indicate that if this were so, the inhabitants' optimum use of the environment would have required that they primarily rely upon pinnipeds from December to May, when mussel, Mytilus californicus, the most abundant mollusk species at the site, toxicity begins to reach dangerous levels (Greengo 1952). Sea bird eggs, stored acorns, and a variety of plants and berries would also have been available at this time (Stodder 1986), as well as spawning fish. Mid-spring and summer months would have provided a variety of small game, sea birds, fish, and plants. These months would also have been profitably spent by hunting deer, elk, and antelope. During late summer and autumn, the inhabitants would have gathered spawning salmon and ripe acorns; shellfish collecting could have resumed shortly before the return of pinnipeds.
Ethnographic Data on Subsistence Behavior
in Coastal California
Due to the lack of available data, most students of Monterey Bay archaeology have encountered difficulties locating ethnographic information on the Costaño tribes. Useful information has been obtained from ethnographies of other California tribes such as the Yuki, Coast Miwok, or southern Pomo (e.g., Kessler 1974; Margolin 1978). Additional information has come from the diaries of explorers and missionaries (Galvin 1964), and from descendents of mission neophytes. It is believed that the Costanoan culture may have closely paralleled that of neighboring tribes. Accounts of subsistence behavior among northern/central coastal Californian Indians suggest that they took full advantage of the abundant animal and plant resources available in the environment.
Fishing
Indians are said to have eaten a wide variety of ocean and freshwater fish, primarily rockfish, smelt, sardines, perch, flounders, rays, lampreys, eels, salmon, and steelhead trout. These fish were caught in a number of different ways. The northern California Tolowa used weirs, nets, and harpoons to obtain freshwater fish; salmon were harpooned from canoes, while smelt were gathered in nets along beaches where they came to spawn on gravel beds (Drucker 1937:232-233). The Coast Yuki, who lacked canoes, and the Coast Yurok, used similar methods, as well as clubs, when dispatching spawning salmon, and throwlines or poles for catching rockfish and trout (Gifford 1939; Kroeber and Barrett 1960). The Coast Miwok near Point Reyes used fish poisons (wild cucumber root) in tide pools at low tide, and are known to have strung nets between two canoes while fishing for surffish. They harpooned salmon and used clubs when they were not selecting for a particular type of fish (Schenk 1970:71-72). According to Margolin (1978), the Costanoans relied heavily upon fishing, and used a variety of nets, weirs, poisons, traps, hooks, and harpoons. Spawning smelt were caught along beaches, while spawning salmon were either captured in nets and clubbed to death, or were lured by bonfires and harpooned along the edges of river banks. Salmon, steelhead, and lampreys appear to have been important to the Costanoans living on the banks of the San Lorenzo and Carmel Rivers (Levy 1978:492).
Once the fish had been caught, they were processed in several ways. Most tribes gutted and salt-washed their catch; it was then split with a stone knife, sun-dried (when intended for winter use), or roasted fresh. Fish were cooked in a number of different ways: some were skewered from head to tail and roasted over a fire, others were hung over the fire, were placed on hot rocks, or were completely buried in hot ashes. Roasted fish was consumed immediately. Lampreys were longitudinally split and sun or smoke dried. The Hupa made a distinction between lampreys which were to be eaten fresh and those which were intended for drying and storage; they were slit with a stone knife or bone awl, respectively. Some groups, such as the Wappo, Karok, and Yurok either pulverized or dried fish vertebrae (a rich source of oil) and saved them for making soup in winter. The Tolowa chose to eat salmon heads, but discarded their vertebrae. The Mattole are known to have fire-hardened salmon ribs-perhaps for use as needles (Kroeber and Barrett 1962; Schenk 1970; Gifford 1939; Driver 1936; Drucker 1937; Leigh 1928).
Interpretation of Fish Remains
from CA-SCR-35
Considering CA-SCR-35's close proximity to fresh and salt water sources, the inhabitants most certainly took advantage of the site's location and fished a great deal. The ethnographic information presented above suggests that there should either be a large amount of burned fish vertebrae in the remains, or practically no vertebrae at all; the latter is the case. The scarcity of fish head bones, and a total number of indeterminate fish vertebrae that account for six fish at the most, suggests that fish were processed in such a way as to prevent preservation of their remains. They were probably pulverized, burned, or boiled.
Bird Resources
Another bountiful resource available to the natives of coastal north-central California was the resident and migratory species of sea birds, waterfowl, inland birds, and birds of prey (for a complete list of local species refer to Gifford and Marshall 1984: Appendix I). The Costanoans, like the Coast Miwok, Wappo, and Yuki tribes, primarily relied upon nets and stuffed bird-skin decoys when hunting waterfowl; Indians spread the nets across streams or hid among the tule reeds and waited for unsuspecting ducks and geese to land (Margolin 1978: Switzer 1978). They paddled tule canoes through reeds and rushes to collect duck eggs, and rowed to off-shore island rookeries to collect gull and cormorant eggs as well as the young chicks. Costanoans are said to have used basket traps, snares, bolas, and strings to capture quail, doves, and other birds (Margolin 1978:25). Birds of prey, especially eagles, buzzards, owls, and ravens were not killed for food.
The Yuki roasted young gulls and cormorants on coals, and the birds' feathers were burned off in the process. They were then gutted, and the heart, liver, kidney, and the gizzard were eaten. Pelicans' fate was to be skinned, eviscerated, and cooked on hot coals. The Costanoans preserved waterfowl by smoking and drying them, and perhaps treated other birds in a similar manner (Gifford 1939:320; Margolin 1978:39). Not all birds' feathers were singed off in the process of cooking--duck and goose feathers provided material for warm cloaks and blankets (Switzer 1974:11; Levy 1978:473). Birds were captured, or their feathers collected, by Coast Miwok girls who sought materials for cloaks and headdresses; particularly prized were those of Redtailed hawks and herons (Schenk 1970:74-77). Whistles and flutes, often elaborately decorated, were fashioned from wing bones (Gifford 1940:161).
Interpretation of Bird Remains
from CA-SCR-35
Birds at CA-SCR-35 are represented mostly by long bone shaft fragments. Processing birds for consumption, according to ethnographic accounts of California Indians, did not include pulverizing as a means of preparation before cooking and eating. It is improbable that aboriginal Californians would have done so, since, when broken, these bone splinters can be quite sharp and rather unpleasant to eat. Also, if the tube-like bones were to have been made into whistles, breaking them would not have been recommended. Whistles, such as those found at CA-ALA-330, were not recognized in the remains; if the inhabitants left them behind on the site, it is quite possible that dairy cow trampling could have broken them together with the majority of the long bones.
In view of its location, it is not surprising that more sea birds and waterfowl than inland birds have been identified from the remains. Many such species, especially gulls, terns, loons, scoters, ducks, and geese, are found nearshore, and feed along beaches or wade in marshes, respectively. The remains, some burned, show that the site's occupants effectively caught and cooked a variety of birds. Burned elements are those of the wings and feet, which have comparatively little flesh on them and would be most likely to burn if roasted. Burned remains of marsh hawk and golden eagle contradict ethnographic accounts of restrictions placed on the consumption of birds of prey. However, two bones are not sufficient evidence to suggest that these birds did not die of natural causes and were subsequently burned. Whether the presence of domestic chicken bones, more than half of which are from the same provenience, among the remains is the result of Costanoan reoccupation of the site during historical times, or of a more recent non-Indian occupation (Gifford and Marshall 1984), has yet to be determined.
Small Game Resources
Small game, particularly gophers, ground squirrels, a variety of rats and mice, skunks, racoons, foxes, coyotes, lynx, and rabbits were also important items in the aboriginal diet. For many tribes, small game provided more food throughout the year than did larger game (Driver 1936:184). Wolves are occasionally reported to have been present in the north-central California coastal environment, and some writers suggest that local tribes successfully hunted them for food and hides (Driver 1936; Margolin 1978). This is highly improbable, since the wolf never reached the western edge of the San Joaquin Valley and was originally most abundant in the north-northeastern region of the state (Mech 1970:31). Trade with more eastern tribes would have been the most probable means by which coastal groups could have procured wolf pelts.
Gophers and ground squirrels were often smoked out of their burrows; other rodents such as woodrats and mice either had their nests burned or were captured in deadfall traps. Skunks and foxes were snared or hit with sticks. These small animals, in addition to rabbits, were gathered from fields after the Indians seasonally burned the shrublands. Rabbits were also caught with throwing sticks, snares, slings, or bow and arrows; they were driven along brush fences where Indians communally captured them in nets or impaled them by thrusting sharp sticks into their burrows (Levy 1978; Margolin 1978; Lewis 1973).
Aboriginal preparation of small game began by roasting an animal over an open fire, unless of course its fur was valued, in which case the animal was undoubtedly skinned before being roasted. Margolin suggests that gopher consumption involved breaking off the animals' tails once the fur had been singed, then burying them in hot ashes directly under the fire (1978:24). It is assumed that meat from small game was roasted and promptly eaten, or else allowed to be dried and smoked for winter rations. Animals whose pelts the Indians particularly esteemed included coyotes, rabbits, and skunks, all of which were made into cloaks; the Wappo made quivers and bags from fox hides (Driver 1936:186). Skunk oil served for medicinal purposes among the Coast Miwok (Schenk 1970:71).
Interpretation of Small Game
Remains from CA-SCR-35
The majority of identifiable mammal remains at CA-SCR-35 are of small game. Note that none of the rodent bones are modified, while slightly larger animals' remains (gray fox and rabbits) are burned (Table 4). Rodents and small animals were almost a staple food for California Indians. Since the Indians were noted for roasting whole rodents--at least gophers--one would expect to find a good deal of burned rodent bones. Recall, however, that only three rodent bones are burned while approximately two dozen very small mammal bones are burned. These data suggest that the site's occupants may have chosen to pulverize Very Small Mammals, as they did fish, thus making it easier to eat rodents without having to remove their bones. How, then, can the lack of burning or heavy damage on the recovered rodent bones be explained?
At least 12 pocket gophers are among the site remains; their bones are in good condition relative to those of other animals, and on two occasions remains of nearly complete gophers were found to have the same provenience. These gophers are thought to have inhabited the site after prehistoric occupation.
Although rabbit remains are abundant (Table 2) at the site, few bones are burned. The majority of burned elements are mandible and maxilla fragments or distal tibias, and no phalanges have been recovered. Since a rabbit's head does not possess much flesh, once its fur is removed the head would be most likely to burn when the animal is roasted. It is apparent that the Indians skinned these rabbits, removing the feet along with the hides, while also leaving the meat on the bone, occasionally exposing articular ends to the fire during roasting, distal tibias in particular.
Remains, some burned, of coyote, fox, skunk, and two larger animals (lynx and puma) are too few for a detailed hypothetical account of activities with specific regard to these animals. One may say that the presence of their remains implies that the site's inhabitants successfully captured and possibly used such animals for meat and furs.
Sea Mammal Resources
Some of the most valuable resources available to the coastal California Indians were sea mammals. In general, the coastal tribes all hunted pinnipeds in a similar manner. Hunters, some disguised with deer or bear skins and charcoal faces, usually paddled canoes to offshore rocks, armed with clubs, harpoons, or bows and arrows (Drucker 1937; Kroeber and Barrett 1960; Gifford 1939; Margolin 1978). A few men disembarked upon reaching the rocks, they slowly approached a sleeping sea lion upwind, often imitating sea lion movements so as not to be discerned or to alarm others. When close enough, they harpooned or clubbed the animal. Harpoons were always aimed at the thorax (Kroeber and Barrett 1960:117); the frontal and parietal/ occipital regions of the cranium are vulnerable areas where sea lions were clubbed. This hunting method is plausible, since sea lions have poor visual acuity when not underwater. They do, however, readily discern rapid movement and sharp, sudden sounds. Such stimuli cause the animals to panic and flee to the safety of open waters (Peterson and Bartholomew 1967:11-12). Southern sea otters, although not members of the pinniped family, northern fur seals, and harbor seals were also approached when sunning or sleeping on rocks and were clubbed or harpooned.
Hunters frequently encountered problems when attempting to retrieve an animal that managed to reach the ocean before they were able to subdue it; one problem was especially common when the animal had been harpooned--it sank (Kroeber and Barrett 1960:116). Euro-American hunters who used rifles and shotguns often experienced this problem when competing against the Alaskan Indians, who were more adept at spear throwing, during the great seal hunts of the 19th century. The Euro-American hunters learned that in order to keep seals from sinking they had to avoid piercing the lungs (Jordan and Clark 1898:143). Aboriginal Californian hunters avoided losing their prey by having one or more men attempt to hold the harpoon line until the animal was exhausted and could be pulled to the surface and speared or clubbed.
After catching the animal, the hunters hoisted it into the canoe, or else punched holes into its flippers and lips so as to tow it ashore. If a seal or sea lion was too large to place in the canoe, the hunters chose whether to butcher it alongside the boat. If they opted to do so, most of the meat, blubber, and skin were taken back to shore (Kroeber and Barrett 1960:119). It is not known if this butchering was done in the water or on the offshore rocks. The latter is more probable because of the impracticality of attempting to dismember a large, non-buoyant animal in deep water. Also, sharks were undoubtedly present in the area and pinniped viscera would have attracted them to the canoe. Smaller individuals were butchered on shore. Pinniped teeth were not saved by Coast Yuki, nor were the animals skinned; the hide was cut up and distributed with the meat. Chunks of hide and meat were placed in fires to singe the hair; later when the fur was gone, the Tolowa washed and cooked them in coals with the fat side down. Meat was sometimes dried, while oil (rendered from fat) was stored in sea lion paunches (Gifford 1939; Drucker 1937; Kroeber and Barrett 1960). Seal and sea lion hides were used for bedding by the Coast Miwok, while most tribes used thick sea otter pelts for blankets or capes.
When whales washed up along the shores, albeit infrequently, their carcasses were butchered and the meat was cut into long, narrow strips which were hung in branches out of bears' reach. The grease was cooked out and saved for later consumption, presumably in sea lion paunches or in baskets (Drucker 1937:237; Margolin 1978:37). The Mattole cracked large bones for marrow; when ribs had been cleaned by the surf they were used as material for making awls and needles and for fires which dried the meat (Kroeber and Barrett 1960:125).
Interpretation of Pinniped and Sea Otter
Remains from CA-SCR-35
With these ethnographic data one may explain several patterns observed within the pinniped remains (Figure 1). Very few vertebrae, ribs, and pelvis fragments are present. Long bones, metapodials, and phalanges, however, are more abundant and are virtually the only elements sustaining evidence of modification. Furthermore, species-identified pinniped long bones are in remarkably good condition. These remains suggest that the CA-SCR-35 occupants eviscerated and butchered pinnipeds away from the site, perhaps so as not to attract scavenging animals. They may also have chosen to reduce the burden to be carried back to camp by selecting only certain portions of a butchered seal or sea lion. Cuts on the proximal humerus suggest that the appendicular portions of these animals had been detached from the carcass. Further sectioning of the limbs, inferred from cuts on the lower limbs, may have occurred at the butchery site or at camp. The rib meat, blubber, and skin were removed along with the choice organs. When butchering pinnipeds at the kill site or on the beach, hunters evidently chose to leave behind the backbone and less often the head. The hunters also appear to have discarded flippers at the butchery site--too few metapodia have been accounted for in view of the minimum number of 13 pinnipeds at the site.
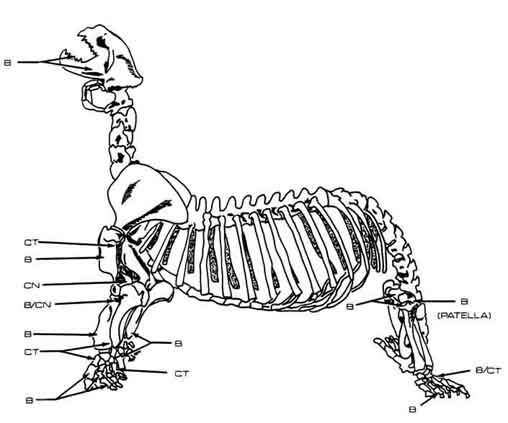
Figure 1. Damage to Bones of All Identified Pinniped Species. B = burned; CT = cut; CN = carnivore gnawed; R = rodent gnawed.
Burned areas on the proximal and distal ends of several long bones suggest that once the limbs had been sectioned the meat was not removed but was instead roasted on the bone over a fire or on hot coals. After the grease had been removed and the meat cooked, it was cut off the bones and either eaten or stored. Many complete, or nearly complete, long bones show that the occupants of CA-SCR-35 did not crack them for marrow. Although the bones are rich in oil, they contain little marrow, and smashing the shafts would not have been worth the trouble.
The five southern sea otters from CA-SCR-35 are represented by long bone and mandible fragments. Even though these animals lack a protective layer of blubber (Woolfenden 1979:22) and are much smaller than pinnipeds, they may have been processed in a similar manner. They were undoubtedly skinned before being roasted, since their pelts were valuable sources of capes and blankets. The total lack of vertebrae, girdle fragments, and most other elements leads one to believe that the animals' limbs were removed from the rest of the body and taken back to camp with the pelt. The limbs were roasted, implied by the fact that all the long bones are quite burned, as are most of the mandible fragments. If the hunters had chosen to take complete animals back to camp, skin them, and roast the carcasses, one would expect to find at least fragmentary remnants of other body parts such as vertebrae or innominates. Even the number of less identifiable Medium Mammal vertebrae do not account for five sea otters. One possible explanation for the discrepancy in element representation is that hunters took whole sea otters back to camp, skinned them, and roasted their limbs, but chose not to strip meat from the rest of the carcass. Instead, the remaining meat was left on the bone, dried, smoked, and stored for later use.
Wild Artiodactyl Resources
Coast mule deer, and to a lesser extent elk and antelope, were undoubtedly the most important game animals hunted by the Costanoans. This fact is documented both archaeologically (e.g., CA-ALA-330 and CA-SMA-22; Phebus 1933:71-73) and ethnographically. Coastal California tribes developed very similar and elaborate rituals the idealized outcome of which was invariably the capture of a deer.
In preparation for the deer hunt, one or more men entered the sweat house--as was the daily procedure--but this time in conjunction with sweating, the hunter(s) abstained from eating meat, fish, salt, and all oily foods; or he (they) may have chosen to fast altogether. In short, a hunter purified himself so as not to be detected by deer which have a well developed olfactory sense (Margolin 1978:31-32; Driver 1936:186; Wallmo 1981:171). He would then rub down with angelica, paint himself with a reddish clay, and don a deer head decoy. The Coast Yuki used a doe's head onto which false antlers were fixed--these were wrapped in fischer skin to resemble velvet (Gifford 1939:316). A similar type of decoy may have been used in the Santa Cruz area; a hunter who attempted to pursue a group of bucks in winter (January-April), when bucks shed their antlers (Wallmo 1981:82), may have been quite successful. During antler shedding season bucks who still possess their antlers dominate those that have lost them (Wallmo 1981:78).
Costanoan hunters most certainly knew where deer would most likely be found. Late fall burning of shrubland and woodland areas produced a succession of grasses, forbs, and browse which increased the potential for successful hunting trips by increasing and improving the amount and quality of available food for large game. This is especially true in late winter-early spring (March-mid April), when available food is normally sparse, and deer are in need of nourishment (Lewis 1973:17-19; Wallmo 1981:351).
Once the herd had been spotted, several stalking strategies were employed by various tribes' hunters. Most commonly, the hunter who wore the decoy began to imitate appropriate deer behavior for that time of the year, while his companions crouched low to remain out of sight and undetected. The decoyed hunter had every interest in letting himself be seen as he approached the herd, because deer escape behavior is triggered by the disappearance of a predator. Deer, as well as elk and antelope, will investigate any new stimulus in their environment, not only out of curiosity, but also to ensure that it poses no immediate or potential threat (Wallmo 1981:172-196; McCullough 1969:71; Nowak 1983:1230). As the deer came within range of the hunters' arrows it was shot.
Another commonly used strategy was for the decoyed hunter to imitate the behavior of the herd, even as it fled in leaps and bounds in fear of his intrusion. Eventually the members, not noticing any strange behavior, would come to accept him. At that time the hunter could select which animal best suited his purposes; he shot it and if lucky, was able to shoot one or two more during the chaos of the herds' escape (Margolin 1978:33). It is improbable that a hunter would have had a large number of animals to choose from. Deer do not normally congregate in large herds; a normal group includes an adult doe, a yearling doe and two fawns of the season. Adult males are found alone or in small groups for most of the year. Larger aggregates of males and females are found during mating season (October-January). However, given the level of aggressiveness and irascibility of the animals at this time of year, a successful hunt would not have been as common as during other seasons. Elk behavior resembles deer's in these respects, antelope however, do form large herds (Nowak 1983:1209-1232). As a rule, the Coast Yuki only shot fawns and bucks, never does (Gifford 1939:316). Those tribes, such as the Tolowa, who depended primarily upon salmon and acorns for subsistence were not known to have used deer head decoys, even though they had heard of them (Drucker 1937:233).
Killing the deer was only half a job done, the hunters had to get the deer back to camp. California tribes within proximity of the coast are known to have done so in one of two ways. The Costanoans are believed to have carried deer carcasses back to their villages, where the animals were skinned, their stomachs were removed and filled with certain organs and choice pieces of meat from "around the kidneys" that were given to a hunter's companions (Margolin 1978:35). The sirloin and other parts of an animal were distributed among villagers, and the brains were placed on a rock to dry for later use in tanning hides (Margolin 1978:35). Wappo hunters, however, butchered their deer at a kill site, as did the Coast Miwok, and divided it equally among themselves; if the hunter was newly married, the deer was taken back to camp and his wife's mother divided it (Driver 1936:185; Schenk 1970:74). A Coast Yuki hunter usually returned to his village after a successful hunt and sent four or five other men to butcher the deer. They butchered the animal with "stone knives" on the ground; it was not suspended by its antlers or hind quarters--a field dressing method preferred by modern sport hunters (Gifford 1939:316; Tinsley 1977:136; Elliot 1975:38-39). Butchered deer meat was wrapped in tanbark oak leaves, tied with hazel twigs and carried back to camp on the men's shoulders. The butchering party occasionally roasted part of the animal at the butchery site. If the kill had been made during the summer, the villagers sometimes "camped out" and feasted on deer meat. The principal hunter was given one hind leg as his share, and the head of the village received the other. Those individuals who received portions of the loin, or back, removed all sinew from the animal and gave it to the hunter, who also had immediate claim to the hide if he chose to take it. The lungs, heart, liver, kidneys, head, and shoulders were anyone's claim, while the fat and meat were particularly valued (Gifford 1939:316-317).
Elk, considerably larger than deer, were butchered as deer were by the Coast Yuki and Tolowa alike. One of two Yuki hunters returned to camp, while the other stayed behind to butcher the animal. During that time the first hunter returned to the kill site with the villagers; they communally cooked the meat and broke the bones for marrow (Gifford 1939:317). The Tolowa chose to partially jerk the animal at the kill site for convenience in carrying the meat back to camp (Drucker 1937:235).
After being brought to camp, fresh meat was always roasted; it was either held over a fire on a stick, laid on coals, or placed in an earth oven. When intended for later use, the meat was cut into strips, and dried either inside or outside the house where it was stored (Drucker 1937:235; Gifford 1939:317; Driver 1936:186).
Coast Miwok hunters backed their bows with sinew, wrapped them in "hide from just above the feet of a deer and glued this on" (Schenk 1970:81). The Coast Yuki felt that only the back sinew was worth saving because it is quite wide and very long. Marrow was occasionally rubbed into wooden bows, presumably to prevent them from drying out and snapping. Hides, an invaluable resource to all aboriginal Californians, were cleaned, de-haired with a rib or shell fragment, dressed with deer brains or with a mixture of brains and oil, and were then worked (Gifford 1939:318; Margolin 1978:30; Drucker 1937:238).
Deer, elk, and antelope were virtually "all-purpose animal resources"; the bones and bone fragments provided material for a variety of tasks. Wappo and Coast Miwok tribes are known to have made detachable harpoon and spear heads from the radius and ulna of deer as well as from deer metapodia, and the Costanoan are said to have used ribs as sweat scrapers (Driver 1936:191; Schenk 1970:72, 82; Margolin 1978:20). Kroeber notes (1962:75) that more northern tribes made similar harpoons from deer "leg bones" and antlers. Apparently,
any fragment of bone which happened to break in such a manner as to produce a sharp point was utilized (to slit lampreys). The first is a mere bone fragment, the naturally sharp point of which has been ground and rounded to some extent. The second is a similarly pointed end of a deer tibia. Sometimes eel splitters are given handles [wooden with cord wrapping] [Kroeber and Barrett 1960:92].
Antlers were saved for awls, wedges, tule saws, and other tools (Margolin 1978:35). The Coast Yuki did not use tule elk bones as material for tools, but found elk antlers quite useful for making spoons and wedges; the Tolowa made them into net gauges and fishing cord shuttles--these tools typically show very smooth, polished and striated surfaces (Gifford 1939:318; Kroeber and Barrett 1960:61). Gifford describes (1939:335) the Coast Yuki process of preparing bones for harpoon barbs: "... longitudinal grooves were cut with giant chiton valves. After cutting half way through, the bone was tapped with a pebble to cause it to split along the grooves."
Morphologically similar bone fragments have been found at several sites (e.g., Emeryville shellmound, Schenck 1926; CA-ALA-330 and CA-SMA-22, Phebus 1973) and many have been designated as tools. Gifford's exhaustive study of Californian bone artifacts (1940) shows that any bone element could have possibly had a functional purpose. However, one must not go upon the assumption that because a given bone, or bone fragment, "looks" like a tool, it necessarily was a tool.
Interpretation of Deer and Elk
Remains from CA-SCR-35
The CA-SCR-35 cervid remains show that the people who occupied the site were successful hunters and were able to benefit from the many resources these animals provided, as did other aboriginal groups.
Margolin's hypothetical anecdote of Costanoan hunters carrying an entire deer back to their village does not appear to have applied to the site's hunters. More likely they killed their deer, gutted it, skinned it, and partially, perhaps fully, butchered it at the kill site. Since escaping intestinal gasses may quickly spoil meat and other internal organs, it would have been more advantageous in terms of edible meat yield if the hunters gutted their deer immediately after slaying it.
One may reasonably infer that an animal has been skinned if cutmarks are present on the shafts and distal shafts of metapodia where little flesh or muscle tissue is present (Frison 1978:314; Binford 1981:107). Such marks are apparent on several mule deer metapodial fragments and first phalanges (Figure 2), elements which may also be modified during the skinning process (Binford 1981:107). Since aboriginal Californians in the region are known to have skinned deer and made use of their hides, it is reasonable to suggest that CA-SCR-35's inhabitants did so as well, and that cuts on these elements are traces of such behavior.
Figure 2. Damage to Bones of Odocoileus hemionus. B = burned; CT = cut; CN = carnivore gnawed; R = rodent gnawed.
Once a deer was gutted and skinned, the CA-SCR-35 hunters opted to butcher it at the kill site. Primary butchery consists of carcass dismemberment in large segments, usually the limbs, ribs, neck and back regions. Examination of deer forelimbs yields very little information about primary butchery. Proximal humeri, which would normally sustain cuts around the rim of the head and on the lateral tuberosity from primary butchering (Binford 1981:123), are completely absent from the assemblage. These elements have a high ratio of spongy, cancellous bone to trabecular bone and are less likely to preserve than other limb elements; thus, they have a low recovery rate. Femurs, whose recovery potential is higher, are more frequently encountered in the remains. The type and location of cuts on proximal femurs suggest that the Costanoans separated the femur from the axial skeleton during primary butchering (Binford 1981:116) by severing the gluteus and biceps muscles and tendon that link it to the pelvis. The Nunamiut Eskimo typically disarticulate the head of the femur from the acetabulum before severing the muscles that link both elements. An immature, cutmarked Odocoileus femur from CA-SCR-35 suggests that the Costanoans did so as well; the femur head (unfused) is present, but the greater trochanter is not. When the animal was dismembered during primary butchering it is quite possible that the unfused greater trochanter remained attached to the pelvis.
The front and hind quarters set aside, the hunters then proceeded to remove the meat from the neck, loin, flank, and ribs (not necessarily in this order); the latter task involved cutting through the sternum and may or may not have involved breaking the ribs in mid-shaft, as it is commonly done by modern butchers (Tinsley 1977:174). The hunters could also have chosen to either break them off near the proximal shaft (Frison 1974:45), or to cut the muscles attaching them to the vertebral column, thus leaving cuts along the proximal shafts (Binford 1981:113). The assemblage's rib shaft fragments are not identifiable beyond the class or order, therefore little evidence is available in support of either method.
Having skinned and butchered their prey, the CA-SCR-35 hunters journeyed back to the village carrying the hide and large butchered segments, perhaps a head and antlers if they were needed. Recall that there are a minimum of seven mule deer at the site; given this number, one would expect to find many vertebrae or vertebrae fragments--such is not the case. Vertebrae, like humeri, have a high ratio of cancellous to compact bone and their potential for survivability in the archaeological record is lower than that of tibias, the MNI determining element. Perhaps Odocoileus vertebrae had been destroyed beyond the possibility of assigning such fragments to the species. To check for their presence in the assemblage, all vertebrae and vertebrae fragments belonging to the indeterminate mammal size, Large Mammal, indeterminate artiodactyl, and Odocoileus sp. were weighed. Collectively, the weight is 276.5 g. When compared to the weight of a complete vertebral column of one adult female mule deer (412.7 g), it is obvious that the CA-SCR-35 hunters did not return to their village carrying an entire deer--only butchered sections previously mentioned and the hide. The frequency of Odocoileus long bones to phalanges demonstrates that the hunters were quite discriminatory about which portions of the animal they chose to carry back to camp. Seven deer should yield a number of phalanges which approximates 125--accounting for the fact that all 168 may not have been preserved or identifiable. A total of 36 out of 40 phalange fragments recovered from the site represent a maximum of three animals. It appears that the majority of the time, when skinning, the hunters chose not to take the phalanges back to camp along with the hides.
Further segmentation of the limbs during secondary butchering may have taken place at the kill site or back at camp; in either case, cutmark patterns show that limbs bearing meat were separated into upper and lower sections. The oblique cuts present on the distal humerus occur when attempting to separate the humerus from the radius and ulna by severing the flexor muscles; this method of dismemberment necessitates that the body be warm and still flexible (Binford 1981:123-124). Such cuts imply one of two possibilities: (a) the deer was completely butchered at the kill site, or (b) it was killed close to camp, where additional butchering was completed. Since few radius and ulna fragments have been recovered from the assemblage, and they possess no cutmarks, it is difficult to tell how the lower forelimbs and main metacarpal were dealt with. Burning patterns on the metapodia, which shall be discussed shortly, suggest that these limb sections were separated at the distal radius and ulna-carpals-proximal metacarpal joint.
Sectioning of the hind limbs at the distal femur and proximal tibia may be inferred from the presence of cuts on the tibial condyles. Disarticulation of the tibia and main metatarsal typically results in cuts along the middle anterior face of the astragalus, a characteristic that is seen on several CA-SCR-35 mule deer astragali (Binford 1981:119-120).
The next step taken by the hunters when processing a mule deer involved one of two procedures: either stripping the flesh off the bones in order to dry the meat, or leaving the meat on the bone and roasting each butchered portion. Burned proximal and distal ends of bones imply that the articular ends were either completely exposed or were not covered with a sufficient amount of meat to prevent burning. The abundance of minimally identifiable rib fragments suggests that ribs were roasted over a fire; meat is only present on one side, and the "bare" side would most likely have been burned. The presence of burned long bone and non-identifiable fragments, the two most abundant modified categories in the assemblage, is explained by the fact that burned bone is structurally weakened and would be expected to break into small pieces.
Meat was cut away from each limb segment either after roasting, or before for filleting. Since little evidence may be obtained from the forelimb remains, this processing activity is inferred from transverse cuts on the proximal anterior and distal posterior femoral shafts (Binford 1981:131-132); similar cuts on proximal tibia shafts, across the crest, also imply such behavior.
Grease and oil were important resources for coastal Californians, so it was to their advantage to crack open the cooked and uncooked limbs and retrieve the marrow; hence the abundance of unmodified long bone shaft fragments and comparatively fewer numbers of identifiable Odocoileus proximal and distal limb fragments.
Ethnographically, elk are known to have been butchered in the same manner as deer. However, it is improbable that the CA-SCR-35 hunters would have chosen to carry more than just the meat back to camp. If it had been the norm to bring back partially butchered segments one would find more elk bone (e.g., articular ends) than is actually present. In all probability, tule elk are less abundant at the site than are mule deer because they are much larger animals, hence their limb segments are heavier. Limb remains belong to immature individuals suggesting that hunters were more willing to transport larger segments of the younger animals. The few vertebrae may have been transported together with the rib and neck meat. One cut thoracic spine implies that the Costanoans severed the tenderloin muscles and segmented the vertebrae during butchering (Binford 1981:111-12) (Figure 3). It is assumed that further elk processing activities paralleled those of mule deer.
Since very few pronghorn antelope remains are present in the assemblage, it is difficult to determine how the CA-SCR-35 inhabitants made use of them. At most one may say that the animals were present in the environment, the Costanoans were equipped to hunt them, and it is probable that the site's inhabitants did.
Figure 3. Damage to Bones of Cervus nannodes. B = burned; CT = cut; CN = carnivore gnawed; R = rodent gnawed.
Artiodactyl Bone Tools
As we have seen, aboriginal Californians made functional tools from bone fragments; assemblage remains show that the CA-SCR-35 inhabitants did so as well (Figures 4, 5, 6). Identified tools resemble several types described by Gifford (1940), and most were described in detail in the previous analysis (Gifford and Marshall 1984). Two are mammal long bone shaft fragments, two are mule deer metapodial shaft fragments, and the remaining two are respectively, a proximal Odocoileus tibia fragment and a proximal metapodial shaft belonging to tule elk. All possessed surface polish and/or striations.
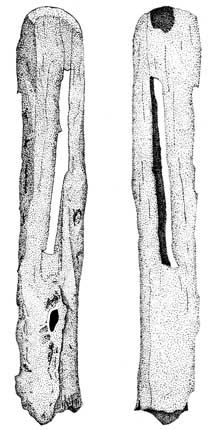

Figure 4. Long Bone Tool Fragments. Left: A. #102420 (length about 8.3 cm). Right: B. #100839 (length about 2.7 cm). Illustrations by Wendy Johnston.
The smallest long bone tool fragment is burned, very scratched, and has a blunt point (Figure 4b). The other (Figure 4a), has a high glossy polish and very smooth edges; at the smoother end the compact bone is worn down and back; the "window break" in mid-shaft is recent.

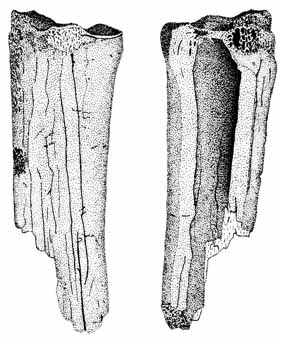
Figure 5. Metapodial Tool Fragments. Left: A. #100914 (length about 8.9 cm). Right: B. #103386 (length about 6.1 cm). Illustrations by Wendy Johnston.
Deep striations and high polish on the tool in Figure 5a suggest that it was not used as a sweat strigil, but was more likely used against a more abrasive surface than human skin--deer hides perhaps. The next tool (Figure 5b) is split down the long axis of the bone; however, the break in mid-shaft is recent.
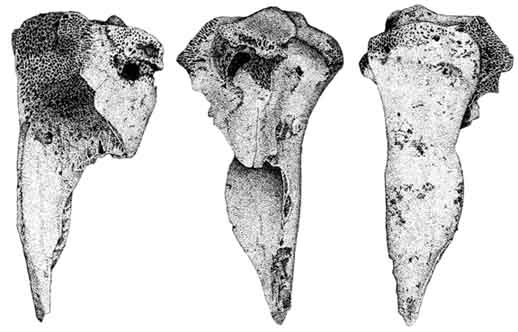
Figure 6a. Mule Deer Tibia Tool Fragment, #101498 (length about 6.6 cm). Illustration by Wendy Johnston.
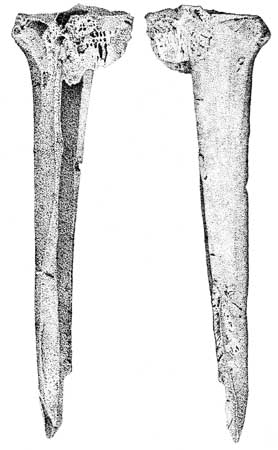
Figure 6b. Tule Elk Metapodial Tool Fragment, #101465 (length about 9.0 cm). Illustration by Wendy Johnston.
The tool in Figure 5b somewhat resembles the elk metapodial (Figure 6a) which has also been longitudinally split, perhaps done in a manner similar to the one practiced by the Coast Yuki. Its broken edges are smooth and polished; one edge has been lightly rodent gnawed. This type of tool most closely resembles awls described by Gifford (1940:161); such tools were used by the Yurok and Hupa as hide punchers and lamprey slitters. Awls appear to have been all-purpose tools for aboriginal Californians. The remaining mule deer tibia (Figure 6b) is highly smoothed and polished, and may also have been used as an eel slitter (Kroeber and Barrett 1960:92).
Domestic Cattle Remains
The presence of domestic cattle remains at the site is not surprising in view of the Eagle Glen dairy farm; both meat bearing and non-meat bearing elements are modified (Figure 7), not necessarily, however, by the Costanoans. Rib fragments are sawed, indicating that they were historically modified, since the Spanish first introduced the practice of saw-butchering cattle (Gust 1982). The distal metapodial, slightly burned, may have sustained several cuts during the removal of its hide. It has also been suggested (Gifford and Marshall 1984) that mission neophytes may have reoccupied the old ancestral site, and were responsible for creating these remains.
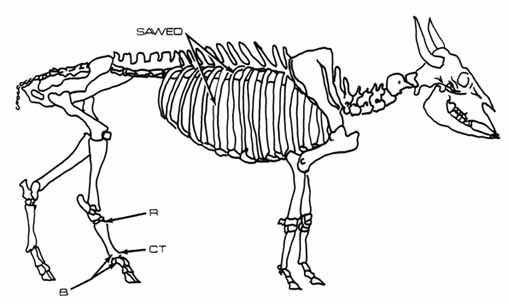
Figure 7. Damage to Bones of Domestic Cattle. B = burned; CT = cut; CN = carnivore gnawed; R = rodent gnawed.
As seen from the presence of their remains, either sheep or goats were on the site-- although there is no other record of this. With the exception of a lumbar vertebra fragment, the remains do not represent meat bearing elements and are not modified, implying that perhaps an entire animal was not introduced at the site, but that a portion of it was consumed there.
California Grizzly Bears as Resources
The largest of all land mammals present in abundance in the Santa Cruz mountains was the California grizzly bear. Aboriginal hunters had ample opportunity to encounter grizzlies, for the animals foraged for berries, acorns, wild grasses, and clover in the hills and grasslands, and are known to have patrolled the beaches in search of marine mammals washed ashore (Storer and Tevis 1955:22-23). Most aboriginal hunters do not appear to have actively sought out grizzlies, but rather avoided them when possible. Costanoans, however, dug pitfall traps over which were suspended pieces of meat. When a grizzly fell into the pit it was dispatched with clubs and bows and arrows (Storer and Tevis 1955:85-87). Grizzlies were such formidable animals that if a Costanoan killed one, he wore a grizzly "tusk" (canine) pendant (Storer and Tevis 1955:88). When they encountered one, Wappo hunters killed a grizzly with bows and arrows or spears; evidently its hide was more valuable than its meat (Driver 1936:186). The Salinans apparently savored grizzly cub meat as opposed to adults' meat (Storer and Tevis 1955:87).
Interpretation of Grizzly Remains
from CA-SCR-35
In view of how large land mammals were butchered, it is not surprising that grizzly bear remains are not present in abundance in the assemblage. Although no ethnographic accounts of grizzly butchering were available, it is likely that the hide and meat were stripped from a bear's carcass--leaving the bones at the kill site. Interestingly, only a complete mandible (an element that possess relatively little meat) represents the species in the assemblage. Speculatively, perhaps the Costanoans held a certain reverence for those who killed grizzlies. A successful hunter may have brought back the hide, meat, and mandible with teeth.
Modifications on the grizzly mandible found at CA-SCR-35 suggest that the animal was killed for its meat as well as its hide. Cutmarks on the element characteristically occur when the masseteric muscle is filleted and when the mandible is loosened so that the tongue may be removed (Binford 1981:109). Taking into account the amount of wear on the grizzly's teeth, especially in view of its omnivorous diet, the animal appears to have been aged at its time of death. Since there may have been an aboriginal preference for younger grizzly meat, this individual may have been killed primarily for its hide.
Bone Distribution at the Site
and Possible Activity Areas
Based on cultural materials recovered from the excavations, the 1979 analysis concluded that CA-SCR-35 was a residential site (Gifford and Marshall 1984). As we have seen, CA-SCR-35 is internally mixed; however, it may still have retained its horizontal integrity (Villa 1982:279). Certain activities, which take place on residential sites, have a tendency to repeatedly occur in specific lateral areas of the site, though are not always restricted to these areas (Binford 1981:310-319). Thus, I plotted the location of each important animal taxon and the types of modifications their remains sustained on plan maps of the site to see how different animals' remains were distributed at CA-SCR-35. Results indicate that the site clearly lacks horizontal integrity. Bird and fish species appear to be concentrated in the Trench 1-3 area; however, there are no well defined activity loci and most of the material, both modified and unmodified, is concentrated in the middle of the site.
SUMMARY AND CONCLUSIONS
We have examined the vertebrate assemblage from CA-SCR-35 as a whole. It is a representative sample of most species prehistorically found in the region. Recent research in the field of taphonomy has enabled us to demonstrate that these faunal remains have been modified by numerous agents. Those agents include chemical and physical weathering, rodents, carnivores, domestic animals, and the prehistoric inhabitants of the Edwards Beach site.
The degree to which chemical and physical weathering has affected the remains is believed to be of little importance relative to other destructive agents. Rodent gnawing is concentrated on harder artiodactyl bone, which is more effective at keeping incisor growth in check. Carnivore gnawing, more prevalent on pinniped remains, is the least common type of animal modification encountered on the remains; in all probability, this is due to limited carnivore access to other animals' remains as a result of human occupation of the site.
Ethnographic accounts of subsistence behavior among neighboring tribes, in conjunction with the data derived from the modified bone assemblage, has allowed a reconstruction of Costanoan subsistence behavior with respect to the animal species found at the site. The CA-SCR-35 Costanoans gathered, snared, and hunted fish, immature and adult birds (marine and freshwater), a variety of small game, from rodents to coyotes, and larger game including pinnipeds, artiodactyls, and an occasional grizzly.
Fish were either gathered in nets or were harpooned and taken back to camp. The small number of species-indentifiable fish remains has been explained as resulting from the ways in which fish were processed: they are believed to have been pulverized, roasted, and perhaps boiled.
Immature sea birds and eggs would most likely have been collected by paddling canoes to nearby offshore rocks. Nets would have been most effective in capturing adult sea birds and various species of migratory and resident waterfowl. Once captured and killed the birds were taken back to camp, plucked and roasted, as implied by the burned bird remains. Burned elements belonging to birds of prey suggest that they may have been consumed by the inhabitants; their feathers could have been of ornamental value.
Small game which comprises the majority of species identified from the assemblage was gathered from burned fields, snared, or trapped. It is reasonable to assume that rodents were a readily available source of food and that they may have been a staple in the aboriginal diet. The lack of identifiable burned rodent bones lead one to believe that they were pulverized and then roasted and eaten. Rabbits and slightly larger animals were undoubtedly skinned for their fur, then roasted (as implied from their burned remains) and consumed, or smoked and dried.
Hunters in tule canoes sought pinnipeds (and southern sea otters) near offshore rocks. They clubbed and harpooned the animals, then brought them ashore to butcher them on the beach; whole sea otters may have been taken back to camp. It is also possible that the hunters chose to butcher larger seals and sea lions at sea. Represented elements, primarily forelimbs, ascertain that butchered segments were transported back to camp, perhaps to be redistributed. Processing would necessarily have required that the grease be removed before the meat. Burned portions of limbs show that meat was roasted while still on the bone; it may also have been dried for storage. Many complete limbs imply that they were not cracked for marrow--in all probability because there is very little to be obtained.
Hunters were also quite successful at capturing wild artiodactyla, primarily deer; this success may in part be attributed to a deer-head decoy or other type of disguise worn by the hunters. Element representation within the cervid assemblage demonstrates that hunters opted to butcher their deer, as well as elk, at the kill site. There, they eviscerated, skinned, and cut up a carcass into more manageable pieces, preparing it for transport back to camp. Thus, the hide and butchered segments were brought to the site, where further butchering was completed if needed. Meat was roasted on the bone; it may also have been smoked and dried for storage. When compared to the condition of pinniped long bones, those of artiodactyls are poorly preserved. The large number of burned and unburned long bone shaft fragments and nonidentifiable bone fragments imply that Indians cracked wild artiodactyl limbs for marrow. Tools were then fashioned from the broken limb fragments.
The grizzly bear, a formidable opponent, apparently was not actively sought by many aboriginal groups; the CA-SCR-35 inhabitants, however, occasionally made use of its hide and its meat.
Results from the analysis of the CA-SCR-35 vertebrate remains demonstrate that the Costanoans share certain subsistence traits with other indigenous California groups. I hope this analysis has not only elucidated possible cultural similarities between the Costanoan inhabitants of CA-SCR-35 and other indigenous tribes, but also stimulated further detailed analyses of faunal assemblages from prehistoric California sites.
END NOTES
* Prepared as a Senior Thesis, University of California, Santa Cruz, June, 1985.
** "Ohlone," which may mean "people of the west" in Miwok, and which may originally have referred to a single group of Costanoan speakers along the San Mateo coast, has been chosen by the Costanoan's descendants as their self-referent term. Thus, in this paper, the term "Costanoan" will be used to describe the prehistoric Indians of the San Francisco-Monterey Bay areas, while "Ohlone" will be used for the modern descendants of that group.
REFERENCES
Allen, J.A. 1974. History of North American Pinnipeds. Arno Press, New York.
Baumhoff, M.A. 1963. Ecological Determinants of Aboriginal California Populations. University of California Publications in American Archaeology and Ethnology 49(2):155-236.
Beals, R. and J. Hester. 1974. California Indians VI. Garland, New York.
Behrensmeyer, A.K. 1978. Taphonomic and Ecologic Information from Bone Weathering. Paleobiology 1(2):150-162.
Binford, L.R. 1981. Bones: Ancient Men and Modern Myths. Academic Press, New York.
Binford, L.R. 1983. Dimensional Analysis of Behavior and Site Structure: Learning from an Eskimo Hunting Stand. In Working at Archaeology, pp. 287-324. Academic Press, New York.
Brown, C.L. and C.E. Gustafson. 1979. A Key to Postcranial Skeletal Remains of Cattle/Bison, Elk, and Horse. Reports of Investigations 57. Washington State University, Pullman.
Burt, W. 1971. Antarctic Pinnipedia. Antarctic Research Series 18(1). American Geophysical Union.
Center for Coastal Marine Studies. 1980. Marine Mammal and Sea Bird Study--Central and Northern California Six Month Progress Report. California Coastal Marine Studies--University of California Santa Cruz. U.S. Department of the Interior, Bureau of Land Management.
Driver, H.E. 1936. Wappo Ethnography. University of California Publications in American Archaeology and Ethnology 36(3):179-220. Berkeley.
Drucker, P. 1937. The Tolowa and their Southwest Oregon Kin. University of California Publications in American Archaeology and Ethnology 36(4):221-300. Berkeley.
Eschmeyer, W., et al. 1983. A Field Guide to Pacific Coast Fishes of North America. Houghton-Mifflin, Boston.
Elliot, C. 1975. Care of Game Meat and Trophies. Funk and Wagnals, New York.
Evans, J. 1978. An Introduction to Environmental Archaeology. Cornell University Press, Ithica.
Frison, G. 1974. The Casper Site: A Hell Gap Bison Kill on the High Plains. Academic Press, New York.
Frison, G. 1978. Prehistoric Hunters of the High Plains. Academic Press, New York.
Galvin, J. (editor). 1964. A Journal of Explorations Northward Along the Coast from Monterey in the Year 1775--Fray Miguel de la Campa. J. Howell, San Francisco.
Gifford, D.P. 1981. Taphonomy and Paleoecology: A Critical Review of Archaeology's Sister Disciplines. In Advances in Archaeological Method and Theory 4. Academic Press, New York.
Gifford, D.P. and D. Crader 1977. A Computer Coding System for Archaeological Faunal Remains. American Antiquity 42:225-238.
Gifford, D.P. and F. Marshall (editors). 1984. Analysis of the Archaeological Assemblage from CA-SCR-35, Santa Cruz County, California. Coyote Press Archives of California Prehistory 2. Salinas.
Gifford, E.W. 1939. The Coast Yuki. Anthropos 34:292-375.
Gifford, E.W. 1940. Californian Bone Artifacts. University of California Anthropological Records 3(2). Berkeley.
Gilbert, B.M. 1980. Mammalian Osteology. Gilbert, Laramie.
Gordon, B. 1979. Monterey Bay Area: Natural History and Cultural Imprints. Boxwood Press, Pacific Grove.
Grayson, D. 1978. On the Quantification of Vertebrate Archaeofaunas. In Advances in Archaeological Method and Theory 2:199-233. Academic Press, New York.
Greengo, R. 1952. Shellfish Foods of the California Indians. Kroeber Anthropology Society Papers 7:63-114.
Gust, S.M. 1982. Faunal Analysis and Butchering. Pp. 101-144 in Ontiveros Adobe: Early Rancho Life in Alto California, J.D. Frierman, ed. Submitted to Redevelopment Agency, Santa Fe Springs.
Hafez, E. et al. 1962. The Behavior of Swine. In The Behavior of Domestic Animals. Williams and Wilkins, Baltimore.
Halley, D. 1980. Chemical and Physical Analysis of Midden Soil at SCR-20, A Coastal Midden Site. Ms. on file, University of California, Santa Cruz, Anthropology Archives.
Harden Jones, F.R. 1968. Fish Migration. Edward Arnold Pub. Co., London.
Harrison, E. 1892. History of Santa Cruz County. Pacific Press, Santa Cruz.
Haynes, G. 1980. Evidence of Carnivore Gnawing on Pleistocene and Recent Mammalian Bones. Paleobiology 6(3):341-351.
Haynes, G. 1983. Frequencies of Spiral and Green Bone Fracture on Ungulate Limb Bones in Modern Surface Assemblages. American Antiquity 48(1):102-114.
Herald, E. 1972. Fishes of North America. Doubleday, New York.
Ingles, L. 1965. Mammals of the Pacific States. Stanford University Press, Stanford.
Jordan, D. and G. Clark. 1898. The History, Needs, and Condition of Fur Seals Resorting to the Pribilof Islands. The Fur Seals and Fur Seals Island of the Northern Pacific Ocean, Part 1, pp. 1-248. Government Printing Office, Washington D.C.
Kessler, C. 1974. Ohlone: Native Americans of the San Francisco and Monterey Bay Areas. Anthropology Senior Thesis, University of California, Santa Cruz.
Kerr, R.M. 1979. Mule Deer Habitat Guidelines. U.S. Department of the Interior, Bureau of Land Management, Denver.
Kroeber, A.L. and S. Barrett. 1960. Fishing Among the Indians of Northwest California. University of California Anthropological Records 21(1):1-210. Berkeley.
Leigh, R. 1928. Dental Pathology of Aboriginal California. University of California Publications in American Archaeology and Ethnology 23(3):399-440. Berkeley.
Levy, R. 1978. Costanoan. In Handbook of North American Indians, Vol. 8, California, pp. 485-495. Smithsonian Institution, Washington, D.C.
Lewall, E. and I. Cowan. 1963. Age Determination in Blacktailed Deer by Degree of Ossification of Epiphyseal Plate in the long Bones. Canadian Journal of Zoology 41(4):629-636.
Lewis, H.T. 1973. Patterns of Indian Burning in California: Ecology and Ethnohistory. Ballena Press Anthropological Papers 1.
Linsdale, J. and P. Tomich. 1953. A Herd of Mule Deer. University of California Press, Berkeley.
Margolin, M. 1978. The Ohlone Way. Heyday Books, Berkeley.
McCullough, D. 1969. The Tule Elk: Its History, Behavior, and Ecology. University of California Press, Berkeley.
McLean, D. 1936. The Replacement of Teeth in Deer as a Means of Age Determination. California Fish and Game 22(1):43-44.
Mech, D.L. 1970. The Wolf--The Ecology and Behavior of an Endangered Species. University of Minnesota Press, Minneapolis.
Monks, G. 1981. Seasonality Studies. In Advances in Archaeological Method and Theory 4:177-240. Academic Press, New York.
Nowak, R. et al. 1983 Walker's Mammals of the World, Vol. 2. 4th ed. John's Hopkins University Press, Baltimore.
Origer, T. 1984. Obsidian Hydration Data. In Analysis of the Archaeological Assemblage from CA-SCR-35, Santa Cruz County, California, D.P. Gifford and F. Marshal, eds. Coyote Press Archives of California Prehistory 2:75-78. Salinas.
Phebus, G. 1973. Contributions to Costanoan Archaeology: Archaeological Investigations at 4-ALA-330 and 4-SMA-22. In Contra Costa Costanoan Archaeology: Archaeological Investigations at 4-ALA-330 and 4-SMA-22. Treganza Anthropology Museum Papers 12. San Francisco State University.
Peterson, R. and G. Bartholomew. 1967. The Natural History and Behavior of the California Sea Lion. Special Publication of the American Society of Mammologists 1. Oklahoma.
Potts, R. and P. Shipman. 1981. Cutmarks Made by Stone Tools on Bones from Olduvai Gorge, Tanzania. Nature 291(5816):577-580.
Rudd, R. 1948. Mammals of Santa Cruz County, California. Master's thesis, University of California, Berkeley.
Schenck, W.E. 1926. The Emeryville Shellmound: Final Report. University of California Publications in American Archaeology and Ethnology 23:147-282.
Schenk, R. 1970. Contributions to the Archaeology of Point Reyes Peninsula: A Compendium in Honor of Ad�n E. Treganza. Treganza Anthropology Museum Papers 6. San Francisco State University.
Shipman, P. 1981. Applications of Scanning Electron Microscopy to Taphonomic Problems. In Annals of New York Academy of Sciences.
Starks, E. 1922. Records of Capture of Fur Seals on Land in California. California Fish and Game 8:155-160.
Stodder, A.L.W. 1986. Mechanisms and Trends in the Decline of the Costanoan Indian Population of Central California. Coyote Press Archives of California Prehistory 4. Salinas.
Storer, T.I. and L.P. Tevis, Jr. 1955. California Grizzly. University of Nebraska Press, Lincoln.
Switzer, L. 1974. An Ethnohistory of the Costanoans of San Francisco Bay. In The Costanoan Indians, R.F. Heizer, ed. Local History Studies, 18. California History Center, DeAnza College, Cupertino.
Tesch, F. 1977. The Eel--Biology and Management of Antuilid Eels. Chapman and Hall, London.
Tinsley, R. 1977. Hunting the Whitetail Deer. Funk and Wagnals, New York.
Villa, P. 1982. Cojoinable Pieces and Site Formation Processes. American Antiquity 47(2):276-287.
Wallmo, O. 1981. Mule and Blacktailed Deer of North America. University of Nebraska Press, Lincoln.
Weaton, H. and V. Brown. 1965. Handbook of California Birds. Naturegraph, Healdsberg.
Wildesen, L. 1969. Notes of an Archaeologist: Ohlone Indian Prehistory. The Indian Historian 2:25-28. San Francisco.
Woolfenden, J. 1979. The California Sea Otter: Saved or Doomed? Boxwood Press, Pacific Grove.
Appendix 1. Weights of Various Taxa.
| Taxon | Weight (g) |
| Vertebrate indet. | 23.9 |
| Mammal indet. | 1,412.1 |
| Very Small Mammal | 28.2 |
| Small Mammal | 103.7 |
| Medium Mammal | 342.9 |
| Large Mammal | 1,551.2 |
| Very Large Mammal | 351.1 |
| Very Very Large Mammal | 134.9 |
| Scapanus latimanus | 0.7 |
| Homo sapiens sapiens | 25.9 |
| Carnivore indet. | 30.9 |
| Small carnivore | 1.2 |
| Medium carnivore | 4.3 |
| Large carnivore | 12.7 |
| Canid | 11.4 |
| Canis sp. | 27.8 |
| Canis latrans | 0.7 |
| Vulpes vulpes | 0.6 |
| Urocyon cinereoargentus | 1.3 |
| Felid | 0.7 |
| Felis sp. | 7.5 |
| Felis concolor | 7.8 |
| Lynx rufus | 1.6 |
| Mustelid | 1.7 |
| Enhydra lutris | 51.2 |
| Gulo sp. | 4.2 |
| Mephitis mephitis | 2.8 |
| Ursus sp. | 6.6 |
| Ursus arctos | 321.6 |
| Pinniped indet. | 53.5 |
| Otarid | 200.0 |
| Eumetopias jubatus | 68.2 |
| Zalophus californianus | 170.3 |
| Arctocephaline | 162.2 |
| Callorhinus ursinus | 217.3 |
| Phoca sp. | 36.5 |
| Phoca vitulina | 23.9 |
| Artiodactyla indet. | 82.0 |
| Medium artiodactyla | 6.6 |
| Medium-large artiodactyla | 150.3 |
| Large artiodactyla | 82.4 |
| Cervus elephus nannodes | 372.3 |
| Odocoileus hemionus | 661.6 |
| Antilocapra americana | 46.0 |
| Bos taurus | 115.7 |
| Caprine indet. | 8.2 |
| Lagenorhyncus obliquidens | 13.3 |
| Rodentia indet. | 23.6 |
| Very small rodent | 1.9 |
| Small rodent | 4.6 |
| Medium rodent | 11.2 |
| Medium-large rodent | 1.5 |
| Sciurus niger | 0.4 |
| Spermophilus beecheyi | 0.5 |
| Neotoma fuscipes | 0.1 |
| Peromyscus sp. | 0.1 |
| Microtus californicus | 2.0 |
| Geomyd | 18.6 |
| Thomomys bottae | 25.7 |
| Leporid | 3.8 |
| Lepus californicus | 9.1 |
| Sylvilagus sp. | 13.6 |
| Sylvilagus bachmani | 6.5 |
| Aves indet. | 59.2 |
| Small aves | 3.5 |
| Medium aves | 13.7 |
| Medium-large aves | 5.0 |
| Gallus sp. | 39.6 |
| Gallus domesticus | 6.6 |
| Ardeidae | 0.3 |
| Butorides virescens | 0.3 |
| Anatidae | 0.7 |
| Aythya sp. | 0.9 |
| Melanitta sp. | 5.2 |
| Melanitta perspicillata | 0.8 |
| Anas sp. | 2.3 |
| Anserine | 2.6 |
| Chen sp. | 1.1 |
| Branta canadensis | 6.2 |
| Larus sp. | 3.7 |
| Sterna sp. | 4.2 |
| Uria sp. | 2.3 |
| Uria aalge | 1.4 |
| Ptychoramphus aleutica | 0.1 |
| Buteo jamaicensis | 1.2 |
| Circus cyanus | 0.4 |
| Aquila chrysaetos | 1.3 |
| Phalocrocorax sp. | 6.7 |
| Phalocrocorax auritus | 6.0 |
| Gavia sp. | 10.4 |
| Tyto alba | 0.1 |
| Fulmarus sp. | 0.4 |
| Fulmarus glacialis | 2.4 |
| Aechmophorus occidentalis | 0.5 |
| Fish indet. | 36.6 |
| Rhacochilus toxotes | 0.3 |
| Raja sp. | 1.0 |
| Oncorhyncus tshawytsha | 0.1 |
| Sardonops sagax | 1.1 |
| Pleuronectidae (cf. right-eyed flounder) | 1.6 |
| Atherinopsis californiensis | 2.3 |
| Total assemblage weight | 7,296.8 |

